
Review Article
Austin J Biomed Eng. 2014;1(4): 1016.
Potential Therapeutic Approaches for Stroke Using Induced Pluripotent Stem Cells
Valerio LSA1,2 and Sugaya K3*
1Institute for Scientific Research and Technology Services (INDICASAT), City of Knowledge, Republic of Panama.
2Department of Biotechnology, Acharya Nagarjuna University, Guntur, India
3Burnett School of Biomedical Sciences, University of Central Florida, USA.
*Corresponding author: :Sugaya K, Burnett School of Biomedical Sciences, College of Medicine, University of Central Florida, 6900 Lake Nona Boulevard, Orlando, FL, 32827, USA.
Received: July 02, 2014; Accepted: July 31, 2014; Published: Aug 01, 2014
Abstract
Stroke is a principal cause of mortality in most countries and up to date there is no cure or potential therapeutic treatment available for this debilitating condition. Human induced pluripotent stem (hiPS) cells are an important source for autologous cells used in regenerative medicine in order to treat brain damage after a stroke has occurred. Recent studies have suggested that transplanted neural cells derived from hiPS cells have the potential to survive, differentiate and improve motor behavior in stroke animal models. In this review, we discuss the conditions needed to reprogram hiPS cells for clinical trials, the strategies used to differentiate hiPS cells toward neural stem cells (NSC) lineage along with their functionality and morphological characteristics studied in vitro. We further address the features founded in vivo after graft transplantation of these neural cells into stroke animal models.
Keywords: Stroke; Stem Cells; Transplantation; Neural tissue
Abbreviations
hiPS: Human induced pluripotent stem cells; NSC: Neural Stem Cells
Introduction
Stroke is a leading cause of mortality and disability worldwide. About 750,000 people experience a modality of strokes each year, and the cost of maintaining these patients’ reaches in excess of billions just in USA [1]. Stroke is a condition in which the brain of an individual is deprived of oxygen and nutrients because the lack of blood supply, which results in death or loss of neural functions. This event is generated by the rupture of an artery/blood vessel in the hemorrhagic stroke, or by the obstruction of a blood vessel for a clot in the ischemic stroke.
The consequences of the damage depend primarily on the area affected, the magnitude of the lesion, the immune response (which may increase damage due to the release of reactive oxygen species and chemokines that attack healthy neurons) and neurorepair mechanisms. If the patients are affected in their locomotor and/or sensory motor areas, then these patients may experience the loss of well-coordinated movements, speech, remembrance, feeling or even total disability. Occipital lobe lesions may disrupt the ability to see; temporal lobe lesion may affect the ability to hear; parietal lobe lesion may interrupt sensory information; and frontal lobe may affect cognition.
Stroke is a sudden and delayed death disease. It is not clear whether a specific genetic mutation or a combination of mutations are directly responsible for the attack, but a study in twins suggests that genetics may be involved in the development of the disease [2]. Interestingly enough, it has been associated with several genes (i.e., the NINJ2 gene, located in chromosome 12p13) as a high-risk biomarker factors to develop hemorrhagic and ischemic stroke [3]. The risk of developing cerebrovascular disease increases in patients with aneurism (which actually is a direct cause of loss of blood supply), hypertension, obesity, high levels of cholesterol in the blood and smoking [1].
A possible treatment of ischemic stroke is recanalization, with the aim to lysate the clots by surgical intervention. An alternative method to lyse the clots is the tissue Plasminogen Activator (tPA) [4], which is the only accepted drug used to treat this disease in clinics at the present. However, this approach has been limited for few patients under specific conditions as the magnitude of the insult (small areas affected), the time since the ictus has occurred (less than 3 hours) and the age of the patient [5]. In contrast, the hemorrhagic stroke has no drugs available to recover the blood supply lost or to improve behavior in patients, and tPA is contraindicated. Importantly, it is known that patients themselves have improved recovery on their own after stroke, probably due to the combination of recovery mechanisms related to neurogenesis.
The application of stem cells in regenerative medicine for stroke has demonstrated to improve recovery after the insult by neurorepair mechanism. However, the use of embryonic stem (ES) cells and adult stem cells (i.e., neural stem cells, mesenchymal stem cells, among others) has different limitations. First, the methods to obtain the samples, involve the death of embryos (in the case of ES cells) or the extraction may be painful and difficult for patients (i.e., adult stem cells from bone marrow). Second, depending on the adult stem cell source, it may be difficult to differentiate the somatic cells of interest. Third, in cases of allogeneic transplantation, autoimmune rejection can occur afterwards because stem cells will have different patterns of molecules from the patient who will receive the graft. Finally, all the limitations together give rise to discrepancies in the ethical concern for uses in clinics and research [6].
hiPS cells prepared for clinical trials
Somatic cells can be reprogrammed to an induced pluripotent stem (iPS) cells by activating pluripotent-signaling pathways shown in Figure 1 [7,8]. These novel kinds of cells with the potency similar to ES cells, avoid the possibility of cell rejection by the host because they are derived from the patient’s own cells (autologous). The major goal of this approach is to ensure an easy source of pluripotent cells, which have the ability to produce somatic cells, including neuroectodermal progenitors without incurring in ethical issues. Thus, iPS cells are a better choice to use in clinical trials compared to their counterpart ES cells for a pluripotent cell source.
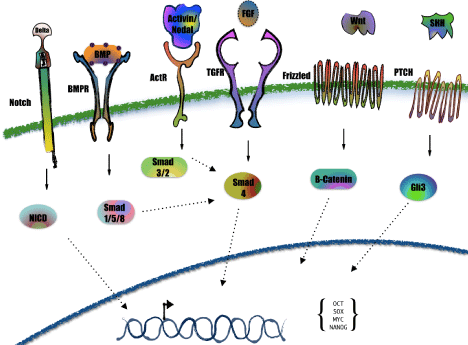
Figure 1: Signaling pathways regulated in pluripotent cells. In this cascades, activation of transcription factors Smad 4, beta catenin and Gli3, regulates the expression of pluripotent factors Oct4, Nanog, Sox2, among others. Bone Morphogenic Proteins (BMP), Activin/Nodal, and Fibroblast Growth Factor (FGF) regulate the Smad 4 protein. Binding of FGF to the FGF receptor (TGFR), directly activates the events which regulates Smad 4. BMP receptor (BMPR) is activated when the ligand BMP binds to the extracellular domain, a process that allows the activation of Smad family members 1, 5 and 8 to regulate Smad 4 activity. Similar occurs with Activin/Nodal ligands attached to the Activin receptors (ActR), but the Smad family members activated are 3 and 2. Wnt protein binds to the receptor Frizzled, a transmembrane protein coupled to G-proteins, to activate beta catenin through multiple phosphorylation events. Sonic Hedgehog (SHH) ligands bind to the PTCH receptor in order to turn on Gli3. The results of these exquisitely coordinated eventsare cells capable of self-renewal and pluripotent capacity, like embryonic stem cells or induced pluripotent stem cells.
There are several reprogramming methods for creating human iPS cells. The first one was the transduction of the Yamanaka Factors (Oct4, c-Myc, Sox2 and k-lif) using a retrovirus [8]. Clinical use of iPS cells produced by this technique has been limited by the random insertion of the transgenes and part of the virus to the genome, the uses of tumorigenic genes such as c-Myc which cause high tumorigenesis, the low rate of successful transfection and the long duration required for reprogramming.
Some of the other technologies used are the episomal vectors [9], the synthetic mRNA [10], the epigenetic regulation of nanog by micro RNAs [11], the piggyBac transposon [12], non-viral plasmid vector [13], and direct delivery of recombinant proteins [14]. These novel technologies have been developed in order to clinically use iPS cells technology, by increasing the efficiency and safety. These technologies allow the production of iPS cells derived from any somatic cell (not limited to fibroblast as the first reprogramming), so that anyone could have access to these iPS cells.
There are some critical considerations for the reprogramming methods to create clinical grade of iPS cells. First, iPS cells need to be free of vectors and transgenes, because transgenes may cause tumorigenesis (i.e., c-Myc). Second, the source of the sample needs to be carefully selected, because sometimes iPS cells suffer spontaneous differentiation (apparently to their somatic ancestor), making it critical to choose the somatic ancestor source to be reprogrammedand derived from the same germ layer tissue to be developed in the future. Third, due to patient needs for therapy, the reprogrammed method needs to be highly efficient in order to generate the best and fastest possible iPS cells. Finally, the iPS cells need to be characterized, demonstrating self-renewal abilities and pluripotent capacity (capable to form endoderm, ectoderm and mesoderm).
In order to bring down the risk of some diseases, it is possible to restore mutations in specific hot spots by creating isogenic controls using iPS cells. Strong evidence in other neurodegenerative diseases, already demonstrates this possibility generating an isogenic control for Parkinson’s disease [15]. If a specific gene were found to be responsible for the development of stroke, then it would be possible to decrease the risk of stroke using this technology. However, more studies need to be evaluated before a practical use for this approach in clinical trials in a safe manner.
Differentiation of hiPS cells toward neural tissue
The ability to produce neural stem cells and their progenitors has been studied deeply in order to understand the neural development and use it for clinical applications as stroke. Figure 2 recapitulates the process used to differentiate hiPS cells toward neural tissue. The objective is to reproduce those neurogenesis events that occur in nature in an in vitro environment. Stroke occurs in organisms that are already developed, meaning adult neurogenesis events should be reproduced, a process that is known to occur in the sub ventricular zone, sub granular zone and dentate gyrus in mammalians. Thus, those brain zones could be possible target areas for cell transplantation for therapy purposes.
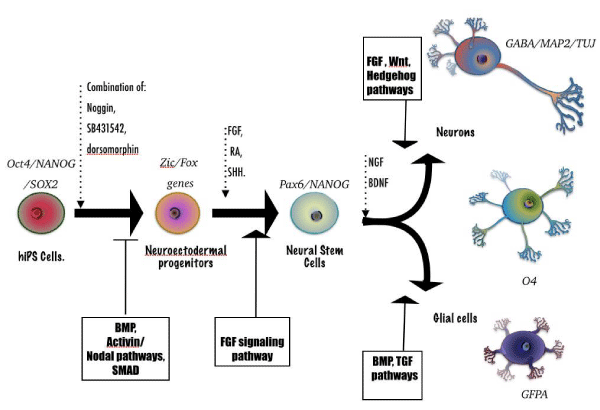
Figure 2: Differentiation of hiPS Cells toward neural tissue. hiPS cells cultures (which express pluripotent factors Oct4, Nanog, Sox2, etc.) are treated with competitors of BMP, Activin and Nodal pathways shown in dot arrows. This leads to the inhibition of Smad signaling. The results are Neuroectodermal Progenitors by activating earlier neuronal genes (i.e., Zic and Fox family members) until the activation of FGF pathway occurs to produce Neural Stem cells (which express neural markers as Nanog and Pax 6). A treatment with terminal neural tissue inducers (i.e., BDNF and NGF) produce the diversity of functional Neurons (expressing TuJ, GABA, MAP2, etc.) and Glial Cells (expressing GFAP, O4, etc.).
The default model for neural ectoderm specification indicates the necessity to block the SMAD signaling through natural inhibition of BMP, Activin and Nodal pathways by Noggin, Follistatin or Chordin (which also occurs artificially by SB431542, dorsomorphin or other competitors of those signals cascades) [16-20]. Apparently, subsequent activation of FGF pathway is needed to activate the right genes involved in the early neural induction process (i.e., Pax, Zic and Fox family genes) [16].
Using this knowledge, several protocols have been developed with the aim to produce in vitro differentiation of pluripotent cells toward neural stem cells (NSC), following the induction of neural tissue for research or therapeutic issues [21-23]. Some of the most used approaches to induce NSC are: the over expression of nanog [24], a combination of retinoic acid (RA) and recombinant sonic hedgehog (SHH), [25], Co-culture of MS5 stromal feeder cells exposed to FGF8 and SHH [26], induction of neural transcription factors [23,27], or by dual inhibition of the SMAD signaling pathway by BMP competitors [18]. These techniques vary in time of differentiation and efficiency.
The combination of nutrients in the culture media (i.e., serum, vitamins, amino acids, etc.) together with the supplemented specific growth factors such as, neuron growth factors (NGF) [28] and brain derived neurotrophic factor (BDNF) [29,30], lead the differentiation of NSC towards terminal neural differentiation. For this process, synergistically activation of canonical Wnt and hedgehog signaling pathways is needed to differentiate NSC into neurons (which express biomarkers as TuJ, GABA and MAP2), whereas BMP signaling pathway activates differentiation to astrocytes (which express biomarkers such as GFAP) and oligodendrocytes (which express biomarkers as O4) [17,31].
Additionally to the growth factors, the scaffold plays an important role for efficient distribution, adhesion and differentiation of the stem cells toward neural tissue. Some novel biomaterials have been developed recently in order to yield neural tissue in a biocompatible manner such as chitosan [32], lactate [33], electrospun poly l-lactic acid/gelatin [34], poly epsilon caprolacton/beta hydroxybutyrate [35], microfibrous [36] and collagen [37]. However, the optimal scaffold is still a challenge and the choice to use one among other depends on economic concerns, reproducibility and whether 2D (monolayers) or 3D cultures (tissue engineering) are needed. In vivo, cells communicate between themselves by autocrine, endocrine and paracrine manner in order to generate functional tissues. In vitro, 3D cultures have the advantage to better mimic the complex niche, morphology and functionality found in neural tissue in vivo, because they grow one layer over another layer forming an extracellular matrix instead of growing in a singular layer. Nevertheless, monolayers have still been useful as an initial step towards producing neural tissue for transplanting or drug screening fields.
Transplantation of NSC derived from hiPS cells into stroke animal models
Recent experiments performed in mice and rats, provided solid evidence that NSC, long-term self-renewing neuroepithelial-like stem cells and neural progenitors derived from iPS cells lines, have the potential to survive, migrate, differentiate and improve recovery after ischemic stroke induced via middle cerebral artery occlusion [20,38-40]. Interestingly, similar results in a rat hemorrhagic stroke model (induced by collagenase) were obtained after transplantation of specific-patient hiPS cells and neural epithelial stem cells derived from hiPS cells [41,42]. In order to study the therapeutic potential of iPS cells in stroke and use it for clinical trials, it is important to take into account the reprogramming method of the iPS cells, the neural differentiation protocol, the transplantation area, the technique for induction of the stroke in the animal model, test if the graft form tumors and the rate of recovery improved. Figure 3 shows the workflow used to study the potential of hiPS cells as a therapy for stroke in animal models.
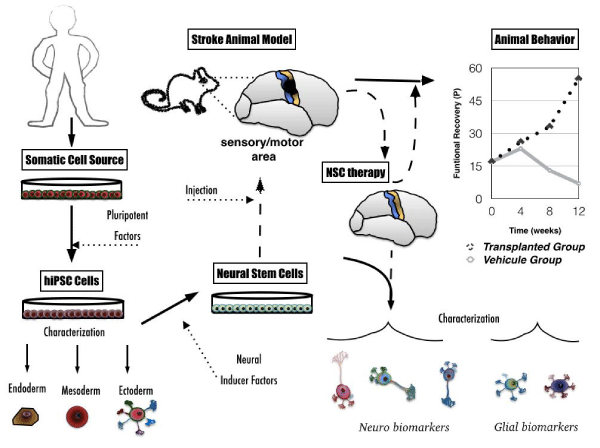
Figure 3: Workflow to evaluate iPS cells as a therapy for stroke in animal models. A culture of somatic cells derived from a biopsy by means of a human donor is needed. Pluripotent factors induce the pluripotent state of the somatic cells (now hiPS cells), making them capable of developing into endoderm, mesoderm and ectoderm. Neural inducer factors lead to the differentiation of hiPS Cells toward NSCs. Transplantation of NSCs grafted into a stroke animal model (i.e., the sensory/motor area affected and low functional recovery index along the time), may improve the functional recovery regardless of the production of the biodiversity of neural cells. Characterization of cells in vitro and post-mortem reveal the biodiversity of functional electrophysiological and morphological neurons and glial cells generated.
Those studies demonstrated in vitro and in vivo, the generation of functional electrophysiological and morphological neurons (i.e., gabaergic, glutamatergic and cholinergic neurons) and glial cells, such as astrocytes and oligodendrocytes. Moreover, it was demonstrated that the graft could establish connections, receive inputs, send axonal projections to different brain layers [39] and improve recovery in the stroke animal models.
The therapeutic methods used for regeneration of brain damage with the aim to improve recovery after ischemic or hemorrhagic stroke focus on redevelopment of the complex microenvironment and mechanisms that have been lost. Whether the immunoreactivity mechanisms produced after the induction of the stroke, or the protocols used to prepare NSC interfere with in vivo differentiation in the animal models as highly efficient as in vitro has seen, is still unclear. Possibly, a better reproducible and controlled manner to generate a stroke animal model, as photothrombosis, could clear this doubt. The transplanted cells respond to cytokines and chemokines released from the host, regardless as to the activation of the immune response, in order to integrate or to reject the graft (i.e., inducing vasculogenesis via VEGF), to migrate to the brain injury (i.e., the chemokine stromal cell-derived factor 1alpha) or to attend the neurorepair mechanism [43,44]. For this reason, the animal model should be chosen carefully to be minimally invasive instead of the more aggressive models used in the pioneer’s studies (i.e., middle cerebral artery occlusion) and more easily reproducible, such as photothrombotic ischemia [45].
The transplantation of 3D cultures of neurons into animal models [36], an approach that should be considered to improve recovery in stroke, was recently demonstrated as being safe. Whether those cells could be safely used in humans is still unclear, specifically due to the high-risk formation of tumorigenesis [38]. So that, more studies need to be evaluated to improve these technologies before using them in clinical trials. Interestingly, it was demonstrated that some compounds such as curcumin could enhance neurogenesis when delivered to animals [46]. This suggests that previous exposure of NSCs to compounds, which enhance cell proliferation and differentiation activity, may increase the rate and velocity of adaptation and biodiversity of the tissue necessary to improve recovery.
Considering the fact that iPS Cells were reprogrammed by using free and unfree vectors and transgenes methods, the different protocols for neuroectodermal specifications cultured in monolayers and the use of different animals models transplanted with grafts at different body areas, these cells have definite shown possibilities to improve brain recovery.
Conclusions
Stroke is one of the principal causes of mortality and disability worldwide. Evidence suggests that genetics may be a determining probability factor to high-risk development for an attack, a fact that is enhanced by other unhealthy influences. hiPS cells are a potential therapeutic treatment for stroke due to their autologous origins eliminating the rejection probabilities, their non-ethical creation, and the fact that they are capable to differentiate into any somatic cell, including NSC. However, hiPS cells for clinical trials still require special considerations in order to maintain their use safely. NSC generated in vitro recapitulates the events occurred in adult neurogenesis. Growth factors, scaffolds and nutrients play an important role in allowing regeneration of the complex microenvironment of neurons. Recent studies strongly make evidenced that the transplantation of neural cells derived from hiPS cells have the potential to survive, to differentiate and to improve behavior in stroke animal models without formation of tumors. Although iPS cells are a potential therapeutic technology for stroke, more studies need to be performed to ensure safety use in clinics.
References
- Go AS, Mozaffarian D, Roger VL, Benjamin EJ, Berry JD, Blaha MJ, et al. Heart disease and stroke statistics-2014 update: a report from the American Heart Association. Circulation. 2014; 129: e28-e292.
- Bak S, Gaist D, Sindrup SH, Skytthe A, Christensen K. Genetic liability in stroke: a long-term follow-up study of Danish twins. See comment in PubMed Commons below Stroke. 2002; 33: 769-774.
- Ikram MA, Seshadri S, Bis JC, Fornage M, DeStefano AL, Aulchenko YS, et al. Genomewide association studies of stroke. See comment in PubMed Commons below N Engl J Med. 2009; 360: 1718-1728.
- del Zoppo GJ, Poeck K, Pessin MS, Wolpert SM, Furlan AJ, Ferbert A, et al. Recombinant tissue plasminogen activator in acute thrombotic and embolic stroke. See comment in PubMed Commons below Ann Neurol. 1992; 32: 78-86.
- Mateen FJ, Nasser M, Spencer BR, Freeman WD, Shuaib A, Demaerschalk BM, et al. Outcomes of intravenous tissue plasminogen activator for acute ischemic stroke in patients aged 90 years or older. See comment in PubMed Commons below Mayo Clin Proc. 2009; 84: 334-338.
- Irons H, Lind JG, Wakade CG, Yu G, Hadman M, Carroll J, et al. Intracerebral xenotransplantation of GFP mouse bone marrow stromal cells in intact and stroke rat brain: graft survival and immunologic response. Cell transplantation. 2004; 13: 283-294.
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. See comment in PubMed Commons below Cell. 2006; 126: 663-676.
- Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. See comment in PubMed Commons below Cell. 2007; 131: 861-872.
- Yu J, Hu K, Smuga-Otto K, Tian S, Stewart R, Slukvin II. Human induced pluripotent stem cells free of vector and transgene sequences. See comment in PubMed Commons below Science. 2009; 324: 797-801.
- Warren L, Manos PD, Ahfeldt T, Loh YH, Li H, Lau F, et al. Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. See comment in PubMed Commons below Cell Stem Cell. 2010; 7: 618-630.
- Lee MR, Prasain N, Chae HD, Kim YJ, Mantel C, Yoder MC, et al. Epigenetic regulation of NANOG by miR-302 cluster-MBD2 completes induced pluripotent stem cell reprogramming. See comment in PubMed Commons below Stem Cells. 2013; 31: 666-681.
- Woltjen K, Michael IP, Mohseni P, Desai R, Mileikovsky M, Hämäläinen R, et al. piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells. See comment in PubMed Commons below Nature. 2009; 458: 766-770.
- Jia F, Wilson KD, Sun N, Gupta DM, Huang M, Li Z, et al. A nonviral minicircle vector for deriving human iPS cells. See comment in PubMed Commons below Nat Methods. 2010; 7: 197-199.
- Zhou H, Wu S, Joo JY, Zhu S, Han DW, Lin T, et al. Generation of induced pluripotent stem cells using recombinant proteins. See comment in PubMed Commons below Cell Stem Cell. 2009; 4: 381-384.
- Soldner F, Laganière J, Cheng AW, Hockemeyer D, Gao Q, Alagappan R, et al. Generation of isogenic pluripotent stem cells differing exclusively at two early onset Parkinson point mutations. See comment in PubMed Commons below Cell. 2011; 146: 318-331.
- Marchal L, Luxardi G, Thomé V, Kodjabachian L. BMP inhibition initiates neural induction via FGF signaling and Zic genes. See comment in PubMed Commons below Proc Natl Acad Sci U S A. 2009; 106: 17437-17442.
- Rajan P, Panchision DM, Newell LF, McKay RD. BMPs signal alternately through a SMAD or FRAP-STAT pathway to regulate fate choice in CNS stem cells. See comment in PubMed Commons below J Cell Biol. 2003; 161: 911-921.
- Chambers SM, Fasano CA, Papapetrou EP, Tomishima M, Sadelain M, Studer L, et al. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. See comment in PubMed Commons below Nat Biotechnol. 2009; 27: 275-280.
- Dincer Z, Piao J, Niu L, Ganat Y, Kriks S, Zimmer B, et al. Specification of functional cranial placode derivatives from human pluripotent stem cells. See comment in PubMed Commons below Cell Rep. 2013; 5: 1387-1402.
- Mohamad O, Drury-Stewart D, Song M, Faulkner B, Chen D, Yu SP, et al. Vector-free and transgene-free human iPS cells differentiate into functional neurons and enhance functional recovery after ischemic stroke in mice. See comment in PubMed Commons below PLoS One. 2013; 8: e64160.
- Kwon J, Lee N, Jeon I, Lee HJ, Do JT, Lee DR, et al. Neuronal Differentiation of a Human Induced Pluripotent Stem Cell Line (FS-1) Derived from Newborn Foreskin Fibroblasts. See comment in PubMed Commons below Int J Stem Cells. 2012; 5: 140-145.
- Swistowski A, Peng J, Liu Q, Mali P, Rao MS, Cheng L, et al. Efficient generation of functional dopaminergic neurons from human induced pluripotent stem cells under defined conditions. See comment in PubMed Commons below Stem Cells. 2010; 28: 1893-1904.
- Panman L, Andersson E, Alekseenko Z, Hedlund E, Kee N, Mong J, et al. Transcription factor-induced lineage selection of stem-cell-derived neural progenitor cells. See comment in PubMed Commons below Cell Stem Cell. 2011; 8: 663-675.
- Sugaya K, Alvarez A, Marutle A, Kwak YD, Choumkina E. Stem cell strategies for Alzheimer's disease therapy. See comment in PubMed Commons below Panminerva Med. 2006; 48: 87-96.
- Hu BY, Zhang SC. Directed differentiation of neural-stem cells and subtype-specific neurons from hESCs. See comment in PubMed Commons below Methods Mol Biol. 2010; 636: 123-137.
- Lee H, Shamy GA, Elkabetz Y, Schofield CM, Harrsion NL, Panagiotakos G, et al. Directed differentiation and transplantation of human embryonic stem cell-derived motoneurons. See comment in PubMed Commons below Stem Cells. 2007; 25: 1931-1939.
- Pang ZP, Yang N, Vierbuchen T, Ostermeier A, Fuentes DR, Yang TQ, et al. Induction of human neuronal cells by defined transcription factors. See comment in PubMed Commons below Nature. 2011; 476: 220-223.
- Kuo YC, Chiu KH. Inverted colloidal crystal scaffolds with laminin-derived peptides for neuronal differentiation of bone marrow stromal cells. See comment in PubMed Commons below Biomaterials. 2011; 32: 819-831.
- Zhang S, Liu XZ, Liu ZL, Wang YM, Hu QL, Ma TZ, et al. Stem cells modified by brain-derived neurotrophic factor to promote stem cells differentiation into neurons and enhance neuromotor function after brain injury. Chinese journal of traumatology = Zhonghuachuangshangzazhi / Chinese Medical Association. 2009; 12: 195-199.
- Kamei N, Tanaka N, Oishi Y, Hamasaki T, Nakanishi K, Sakai N, et al. BDNF, NT-3, and NGF released from transplanted neural progenitor cells promote corticospinal axon growth in organotypic cocultures. See comment in PubMed Commons below Spine (Phila Pa 1976). 2007; 32: 1272-1278.
- Menendez L, Yatskievych TA, Antin PB, Dalton S. Wnt signaling and a Smad pathway blockade direct the differentiation of human pluripotent stem cells to multipotent neural crest cells. Proceedings of the National Academy of Sciences of the United States of America. 2011; 108: 19240-19245.
- Shi W, Nie D, Jin G, Chen W, Xia L, Wu X, et al. BDNF blended chitosan scaffolds for human umbilical cord MSC transplants in traumatic brain injury therapy. See comment in PubMed Commons below Biomaterials. 2012; 33: 3119-3126.
- �lvarez Z, Casta�o O, Castells AA, Mateos-Timoneda MA, Planell JA, Engel E, et al. Neurogenesis and vascularization of the damaged brain using a lactate-releasing biomimetic scaffold. See comment in PubMed Commons below Biomaterials. 2014; 35: 4769-4781.
- Binan L, Tendey C, De Crescenzo G, El Ayoubi R, Ajji A, Jolicoeur M, et al. Differentiation of neuronal stem cells into motor neurons using electrospun poly-L-lactic acid/gelatin scaffold. See comment in PubMed Commons below Biomaterials. 2014; 35: 664-674.
- Kuo YC, Huang MJ. Material-driven differentiation of induced pluripotent stem cells in neuron growth factor-grafted poly(ε-caprolactone)-poly(β-hydroxybutyrate) scaffolds. See comment in PubMed Commons below Biomaterials. 2012; 33: 5672-5682.
- Lu HF, Lim SX, Leong MF, Narayanan K, Toh RP, Gao S, et al. Efficient neuronal differentiation and maturation of human pluripotent stem cells encapsulated in 3D microfibrous scaffolds. See comment in PubMed Commons below Biomaterials. 2012; 33: 9179-9187.
- Guan J, Zhu Z, Zhao RC, Xiao Z, Wu C, Han Q, et al. Transplantation of human mesenchymal stem cells loaded on collagen scaffolds for the treatment of traumatic brain injury in rats. See comment in PubMed Commons below Biomaterials. 2013; 34: 5937-5946.
- Kawai H, Yamashita T, Ohta Y, Deguchi K, Nagotani S, Zhang X, et al. Tridermal tumorigenesis of induced pluripotent stem cells transplanted in ischemic brain. See comment in PubMed Commons below J Cereb Blood Flow Metab. 2010; 30: 1487-1493.
- Oki K. [Stem cell therapy for ischemic stroke using iPS cells]. See comment in PubMed Commons below Rinsho Shinkeigaku. 2013; 53: 1017-1019.
- Jensen MB, Yan H, Krishnaney-Davison R, Al Sawaf A, Zhang SC. Survival and differentiation of transplanted neural stem cells derived from human induced pluripotent stem cells in a rat stroke model. Journal of stroke and cerebrovascular diseases: the official journal of National Stroke Association. 2013; 22: 304-308.
- Qin J, Gong G, Sun S, Qi J, Zhang H, Wang Y, et al. Functional recovery after transplantation of induced pluripotent stem cells in a rat hemorrhagic stroke model. See comment in PubMed Commons below Neurosci Lett. 2013; 554: 70-75.
- Yuan M, Wen SJ, Yang CX, Pang YG, Gao XQ, Liu XQ, et al. Transplantation of neural stem cells overexpressing glial cell line-derived neurotrophic factor enhances Akt and Erk1/2 signaling and neurogenesis in rats after stroke. See comment in PubMed Commons below Chin Med J (Engl). 2013; 126: 1302-1309.
- Lee HJ, Kim KS, Park IH, Kim SU. Human neural stem cells over-expressing VEGF provide neuroprotection, angiogenesis and functional recovery in mouse stroke model. See comment in PubMed Commons below PLoS One. 2007; 2: e156.
- Imitola J, Raddassi K, Park KI, Mueller FJ, Nieto M, Teng YD, et al. Directed migration of neural stem cells to sites of CNS injury by the stromal cell-derived factor 1alpha/CXC chemokine receptor 4 pathway. See comment in PubMed Commons below Proc Natl Acad Sci U S A. 2004; 101: 18117-18122.
- Labat-gest V, Tomasi S. Photothrombotic ischemia: a minimally invasive and reproducible photochemical cortical lesion model for mouse stroke studies. See comment in PubMed Commons below J Vis Exp. 2013.
- Dong S, Zeng Q, Mitchell ES, Xiu J, Duan Y, Li C, et al. Curcumin enhances neurogenesis and cognition in aged rats: implications for transcriptional interactions related to growth and synaptic plasticity. See comment in PubMed Commons below PLoS One. 2012; 7: e31211.