
Review Article
Austin J Musculoskelet Disord. 2016; 3(2): 1035.
Involvement of Lipid and Glucose Metabolism in Skeletal Disorders of Animal Models
Hiroshi Kawaguchi*
The Chief of the Spine Center, Japan Community Health Care Organization (JCHO), Japan
*Corresponding author: Hiroshi Kawaguchi, The Chief of the Spine Center, Japan Community Health Care Organization (JCHO), Tsukudo 5-1, Tokyo 162-8543, Japan
Received: June 06, 2016; Accepted: September 20, 2016; Published: September 22, 2016
Abstract
Degenerative skeletal diosrders are now considered a major public health issue causing chronic disability in most developed countries. This paper summarizes our mouse genetics studies on the molecular backgrounds of representative degenerative skeletal diosrders: osteoporosis, Ossification of the Posterior Longitudinal Ligament of the Spine (OPLL), and bone fracture healing. By analyzing deficient mice, PPARγ, a key adipogenesis molecule intrinsic to bone marrow progenitors, was shown to be involved in age-related osteoporosis. Studies on deficient mice and OPLL patients revealed that insulin and insulin-like growth factor-I (IGF-I) are potent bone anabolic factors through the balance of distinct signals of the two adaptor molecules, Insulin Receptor Substrate (IRS)-1 and IRS-2: IRS-1 for maintenance of bone turnover by up-regulating anabolic and catabolic functions of osteoblasts, while IRS- 2 for retaining the predominance of the anabolic function over the catabolic function. IRS-1 was also essential for bone fracture healing. These molecules could be therapeutic targets for the skeletal disorders. Although we used the mouse genetics approaches, we have attempted to confirm the reproducibility of the mouse findings in humans using human gene polymorphism or clinical biochemical studies. Among the molecules we identified in this study, there are some whose suppression ameliorated skeletal disorders under pathological conditions but did not affect physiological conditions, indicating that targeting on these molecules may lead to an ideal treatment without side effects on physiological functions. In fact, trials based on the present findings are being practically planned for clinical application.
Introduction
Due to the rapidly increasing number of elderly individuals today, degenerative skeletal diosrders are now considered a major public health issue causing chronic disability in most developed countries. Among them, osteoporosis is a representative disorder worldwide with strong social impacts [1]. Ossification of the Posterior Longitudinal Ligament of the Spine (OPLL) with a prevalence of 2-4% in Asia and somewhat lower in other countries is a disorder that causes severe tetra paresis [2]. Despite significant social demand for more information of the etiology of the disorders, the molecular mechanisms have not yet been fully elucidated. The mouse genetics approach is now considered to be a useful and efficient method due to recent progression of gene manupulating techniques in vivo. This manuscript introduces our recent mouse genetics studies attempting to elucidate the molecular mechanisms underlying the bone and cartilage disorders.
PPARγ as an Intracellular Molecule Regulating Age-Related Osteoporosis
Ageing is one of the major causes of osteoporosis, and the underlying mechanisms include intracellular and extracellular signals of osteoblastic cells. As an intracellular molecule regulating agerelated osteoporosis, Runx2, a key transcription factor for osteoblast differentiation, is the most probable candidate, since the expression is reported to be decreased during cellular ageing of osteoblasts in vitro [3]. However, there is no in vivo evidence of its contribution to bone loss with ageing.
Osteoblasts and adipocytes are known to share a common progenitor: multipotential mesenchymal cells in bone marrow, being driven by respective key molecules Runx2 and PPARγ (Figure 1A) [4]. In addition, ageing is associated with a reciprocal decrease of osteogenesis and an increase of adipogenesis in bone marrow [5]. Hence, we investigated the role of PPARγ in bone metabolism by analyzing heterozygous PPARγ-deficient (PPARγ+/–) mice [6,7], since the homozygous deficient (PPARγ–/–) mice were embryonically lethal [8]. Although they showed no abnormality in major organs, PPARγ+/– mice exhibited higher bone mass than the Wild-Type (WT) littermates, especially at older ages (Figure 1B), indicating the involvement of the PPARγ signal in the pathophysiology of agerelated osteoporosis [6,7]. Ex vivo cultures of bone marrow cells showed that PPARγ haplo insufficiency caused not only a decrease in the number of adipocytes, but also an increase of osteoblasts (Figure 1C), indicating that PPARγ signal in marrow progenitors functions as a potent suppressor of commitment to osteoblastic lineage. Clinical involvement of PPARγ in bone loss was confirmed by an association study between the human gene polymorphism and bone density in Japanese postmenopausal women [9] (Figure 1).
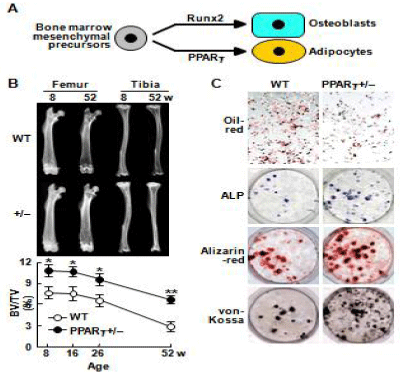
Figure 1: (A) schema of differentiation of osteoblasts and adipocytes. (B)
Radiographs of WT and PPARγ+/– bones at 8 and 52 weeks of age, and time
course of trabecular bone volume (BV/TV) at the distal femora measured on
the CT image. *P<0.05, #P<0.01 vs. WT. (C) Adipogenesis by oil-red staining
and osteogenesis by alkaline-phosphatase (ALP), Alizarin-red, and von-
Kossa stainings in cultures of WT and PPARγ+/– bone marrow cells.
IGF-I/ IRS Signal Regulating Age-related Osteoporosis
There are several cytokines / growth factors that function as extracellular signals regulating age-related osteoporosis [10], and among them IGF-I is the most probable candidate since the serum level is positively correlated with bone density of aged populations [11]. Patients with Laron syndrome caused by IGF-I deficiency also exhibit severe osteoporosis [12], indicating that IGF-I is a potent bone anabolic factor clinically.
IGF-I as well as insulin initiate cellular responses by binding to their respective cell-surface receptors, and then activate essential adaptor molecule Insulin Receptor Substrate (IRS) followed by downstream signaling pathways like phosphatidylinositol-3 kinase (PI3K)/Akt and mitogen-activated protein kinases (MAPKs) [13]. Among the IRS family, we found that IRS-1 and IRS-2 are expressed in bone [14,15]. Our further studies on mice lacking IRS-1 (IRS–1– /–) or IRS-2 (IRS–2–/–) revealed that these mice exhibited severe osteopenia with distinct mechanisms: IRS-1–/– mice showed a low bone turnover in which both bone formation and resorption were decreased (Figure 2) [14]. Whereas IRS-2–/– mice showed an uncoupling status with decreased bone formation and increased bone resorption (Figure 3) [15]. It therefore seems that IRS-1 is important for maintaining bone turnover, while IRS-2 for remodeling balance retaining the predominance of anabolic function over catabolic function of osteoblasts. IGF-I and insulin may up-regulate bone formation without affecting bone resorption through the balance of the two IRS signals in osteoblasts (Figure 4) [16].
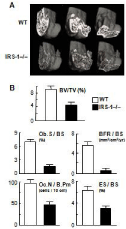
Figure 2: (A) 3D-CT of distal femurs of WT and IRS-1–/– littermates at 8
weeks of age. (B) Bone histomorphometric analysis of the proximal tibiae.
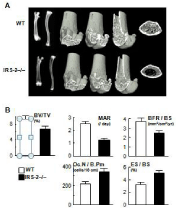
Figure 3: (A) Radiographs of femurs and tibias, and 3D-CT of distal femurs of
WT and IRS-2–/– littermates at 8 weeks of age. (B) Bone histomorphometric
analysis of the proximal tibiae.
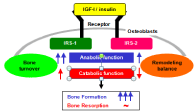
Figure 4: A schema of mechanism underlying the osteoanabolic function of
insulin and IGF-I via distinct functions of IRS-2 and IRS-2 in osteoblasts.
Insulin Signal in OPLL
Patients with type 1 diabetes with decreased insulin level are often associated with osteoporosis [17]. Probably via the suppression of IRS-1 and IRS-2 signals above. Besides osteoporosis, the insulin/ IGF-I signal may possibly be involved in other skeletal disorders. Although the relationship between osteoporosis and type 2 diabetesis controversial, most OPLL patients have been reported to accompany type 2 diabetes [18]. Our clinical study examining the relationship between the extent of ossification and glucose intolerance in OPLL patients revealed that the insulin secretory response, but not the severity of glucose intolerance or diabetes, was associated with the ossification severity (Figure 5), [19,20] confirming the clinical osteoanabolic action of insulin under the pathological condition (Figure 5).
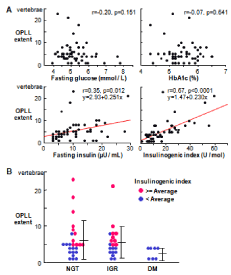
Figure 5: (A) Correlation between OPLL extent and serum levels of diabetesrelated
factors in OPLL patients (n=52). (B) OPLL extent of patients with three
grades of diabetes severity: Non-Glucose Tolerance (NGT), Intermediate
Glucose Resistance (IGR), and Diabetes Mellitus (DM). Patients with
insulinogenic index equal to or higher than the average are shown as closed
circles and those less than that are shown as open circles.
Taking the clinical and mouse genetics studies together, in OPLL patients with type 2 diabetes the up-regulation of insulin production as a feedback mechanism of the impairment of insulin sensitivity in the target cells may stimulate osteoprogenitor cells in the ligament to induce ossification. Contrarily, in IRS-1–/– and IRS-2–/– mice even the increased insulin may not affect osteoblasts, resulting in osteoporosis (Figure 6).
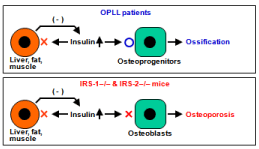
Figure 6: A schema of mechanisms underlying the ossification in OPLL
patients, and osteopenia and impaired fracture healing in IRS-1–/– mice.
IRS-1 Signal in Bone Fracture Healing
To further know the involvement of IRS-1 in bone formation, we compared the healing of bone fracture which was created at the mid shaft of mouse tibia and fixed with an intramedullary nail in IRS-1–/– mice (Figure 7A) [21]. After 3 weeks, all 15 WT mice showed bone union, while in IRS-1–/– mice, only four showed bone union but with small fracture callus, and the other 11 mice remained non-union (Figure 7B & C). Contrarily, the fracture healing was not suppressed in IRS-2 mice (data not shown). In the ex vivo cultures of WT osteoblasts, cell proliferation stimulated by IGF-I was suppressed strongly by an inhibitor of Aktsignal, moderately by an ERK inhibitor, but not by a p38 MAPK inhibitor (Figure 7D). Meanwhile, IRS-1–/– osteoblasts were less respondent to IGF-I and were inhibited only by the ERK inhibitor. These indicate that the osteoanabolic action of IGF-I/IRS-1 is mainly mediated by the Akt pathway.
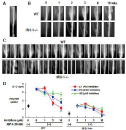
Figure 7: (A) The mouse fracture model. (B) Time course of fracture healing
in representative WT and IRS-1–/– mice. (C) Radiographs of fracture sites
of all WT mice (n=15) and IRS-1–/– mice (n=15) 3 weeks after the fracture.
(D) Proliferation of primary chondrocytes determined by [3H]-TdR uptake by
IGF-I and specific inhibitors. Means (symbols) ± S.E. (error bars) for 8 wells/
group.*p<0.01 vs. IGF-I alone.
Conclusion
The insulin-IGF-I/IRS-1-IRS-2 /Akt signal may play crucial roles for bone formation under various physiological and pathological conditions. It maintains bone volume and turnover for prevention of osteoporosis, enhances OPLL progression, and facilitates bone fracture healing. Further understanding of the molecular network related to this signal will provide a basis for rational therapeutic targets for these disorders.
The ultimate aim of the present study is to identify the molecular targets for clinical treatments of the degenerative skeletal disorders. Although we mainly used the mouse genetics approaches, we have attempted to confirm the reproducibility of the mouse findings in humans using human gene polymorphism or clinical biochemical studies [22-27]. Among the molecules we identified in this study, there are some whose suppression ameliorated skeletal disorders under pathological conditions but did not affect physiological conditions, indicating that targeting on these molecules may lead to an ideal treatment without side effects on physiological functions. In fact, trials based on the present findings are being practically planned for clinical application.
References
- Clark PM, Ellis BM. A public health approach to musculoskeletal health. Best Pract Res ClinRheumatol. 28; 2014: 517-532.
- Abiola R, Rubery P, Mesfin A. Ossification of the posterior longitudinal ligament: Etiology, diagnosis, and outcomes of nonoperative and operative management. Global Spine J: 2016; 195-204.
- Christiansen M, Kveiborg M, Kassem M, Clark BF, Rattan SI. CBFA1 and topoisomerase I mRNA levels decline during cellular aging of human trabecular osteoblasts. J Gerontol A Biol Sci Med Sci. 2000; 55: B194-200.
- Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multiline age potential of adult human mesenchymal stem cells Science. 1999; 284: 143-147.
- Meunier P, Aaron J, Edouard C, Vignon G. Osteoporosis and the replacement of cell populations of the marrow by adipose tissue. A quantitative study of 84 iliac bone biopsies. Clin Orthop. 80: 1971; 147-154.
- Akune T, Ohba S, Kamekura S, Yamaguchi M, Chung U, Kubota N, et al. PPARγ insufficiency enhances osteogenesis through osteoblast formation from bone marrow progenitors. J Clin Invest. 113: 2004; 846-855.
- Kawaguchi H, Akune T, Yamaguchi M, Ohba S, Ogata N, Chung UI, et al. Distinct effects of PPARγ insufficiency on bone marrow cells, osteoblasts, and osteoclastic cells. J Bone Miner Metab. 23: 2005; 275-279.
- Kubota N, Terauchi Y, Miki H, Tamemoto H, Yamauchi T, Komeda K, et al. PPARγ gamma mediates high-fat diet-induced adipocyte hypertrophy and insulin resistance. Mol Cell. 1999; 4: 597-609.
- Ogawa S, Urano T, Hosoi T, Miyao M, Hoshino S, Fujita M, et al. Association of bone mineral density with a polymorphism of the peroxisome proliferator-activated receptor gamma gene: PPARγ gamma expression in osteoblasts. Biochem Biophys Res Commun. 1999; 260: 122-126.
- Harada S, Rodan GA. Control of osteoblast function and regulation of bone mass. Nature. 2003; 423: 349-355.
- Rosen CJ. Growth hormone, insulin-like growth factors, and the senescent skeleton: Ponce de Leon's Fountain revisited? J Cell Biochem. 1994; 56: 348-356.
- Laron Z, Klinger B, Silbergeld A. Patients with Laron syndrome has osteopenia/osteoporosis. J Bone Miner Res:1999; 14: 156-157.
- Kadowaki T, Tobe K, Honda-Yamamoto R, Tamemoto H, Kaburagi Y, Momomura K, et al. Signal transduction mechanism of insulin and insulin-like growth factor-1. Endocr J. 43: 1996; 33-41.
- Ogata N, Chikazu D, Kubota N, Terauchi Y, Tobe T, Azuma Y, et al. Insulin receptor substrate-1 in osteoblast is indispensable for maintaining bone turnover. J Clin Invest. 2000; 105: 935-943.
- Akune T, Hoshi K, Kubota Y, Terauchi K, Tobe Y, Azuma T, et al. Insulin receptor substrate-2 maintains predominance of anabolic function over catabolic function of osteoblasts. J Cell Biol. 2002; 159: 147-156.
- Kawaguchi H. Molecular backgrounds of age-related osteoporosis from mouse genetics approaches. Rev Endocr Metab Disord. 2006; 7: 17-22.
- Thrailkill KM, Lumpkin CK Jr, Bunn RC, Kemp SF, Fowlkes JL. Is insulin an anabolic agent in bone? Dissecting the diabetic bone for clues. Am J Physiol Endocrinol Metab. 2005; 289: 735-745.
- Shingyouchi Y, Nagahama A, Niida M. Ligamentous ossification of the cervical spine in the late middle-aged Japanese men. Its relation to body mass index and glucose metabolism Spine. 1996; 21: 2474-2178.
- Akune T, Ogata N, Seichi A, Ohnishi I, Nakamura K, and Kawaguchi H. Insulin secretory response is positively associated with the extent of ossification of the posterior longitudinal ligament of the spine. J Bone Joint Surg (Am). 2001; 83: 1537-1544.
- Kawaguchi H, Akune T, Ogata N, Seichi A, Takeshita K, and Nakamura K: Contribution of metabolic conditions to ossification of the posterior longitudinal ligament of the spine. In: Yonenobu K, Nakamura K, Toyama Y, editors. OPLL: Ossification of the Posterior Longitudinal Ligament. 2nd ed. Springer. 2006; 37-40.
- Shimoaka T, Kamekura S, Chikuda H, Hoshi K, Chung U, Akune T. Impairment of bone healing by insulin receptor substrate-1 deficiency. J Biol Chem. 2004; 279: 15314-15322.
- Ogata N, Shiraki M, Hosoi T, Koshizuka Y, Nakamura K, and Kawaguchi H. A polymorphic variant at the Werner helicase (wrn) gene is associated with bone density, but not spondylosis, in postmenopausal women. J Bone Miner Metab. 2001; 19: 296-301.
- Koshizuka Y, Kawaguchi H, Ogata N, Ikeda T, Mabuchi A, Seichi A, et al. Nucleotide pyrophosphatase gene polymorphism associated with ossification of the posterior longitudinal ligament of the spine. J Bone Miner Res. 2002; 17: 138-144.
- Ogata N, Matsumura Y, Shiraki M, Kawano K, Koshizuka Y, Hosoi T. Association of klotho gene polymorphism with bone density and spondylosis of the lumbar spine in postmenopausal women. Bone. 2002; 31: 37-42.
- Ogata N, Koshizuka Y, Miura T, Iwasaki M, Hosoi T, Shiraki M, et al. Association of bone metabolism regulatory factor gene polymorphisms with susceptibility to ossification of the posterior longitudinal ligament of the spine and its severity Spine. 2002; 27: 1765-1771.
- Kawano K, Ogata N, Chiano M, Molloy H, Kleyn P, Spector T D, et al. Klotho gene polymorphisms associated with bone density of aged postmenopausal women. J Bone Miner Res. 17: 2002; 1744-1751.
- Koshizuka Y, Ogata N, Shiraki M, Hosoi T, Seichi A, Takeshita K, et al. Distinct association of gene polymorphisms of estrogen receptor and vitamin D receptor with lumbar spondylosis in postmenopausal women. Eur Spine J. 2006; 15: 1521-1528.