
Research Article
Austin J Nutri Food Sci. 2015; 3(4): 1071.
Kinetics of Renin and Angiotensin Converting Enzyme Inhibition by African Giant Land Snail (Archachatina marginata) Protein Hydrolysate and Membrane Ultrafiltration Peptide Fractions
Girgih AT¹, Nwachukwu ID¹, Iwar MI², Fagbemi TN³ and Aluko RE¹*
¹Department of Human Nutritional Sciences, University of Manitoba, Canada
²Department of Wild Life and Range Management, Federal University of Agriculture, Makurdi, Nigeria
³Department of Food Science and Technology, Federal University of Technology, Akure, Nigeria
*Corresponding author: Rotimi E. Aluko, Department of Human Nutritional Sciences, University of Manitoba, Winnipeg, MB R3T 2N2, Canada
Received:August 27, 2015;Accepted:December 04, 2015;Published: December 11, 2015
Abstract
The aim of the study was to produce potential antihypertensive peptides with multi-enzyme inhibitory effects from defatted snail protein meal (SnPM). The SnPM was enzymatically hydrolysed by sequential pepsin and pancreatin addition to mimic gastrointestinal tract digestion. After the digestion, approximately 81% of the initial SnPM protein was converted into soluble peptides, which were collected as the snail protein hydrolysate (SnPH). The SnPH was then fractionated using ultrafiltration membranes to obtain peptide fractions with <1, 1-3, 3-5, 5-10 and >10 kDa molecular weight sizes. The SnPH and fractionated peptides were investigated for in vitro inhibitions of angiotensin I-converting enzyme (ACE) and renin activities followed by determination of enzyme inhibition kinetics parameters.The SnPH had 68 and 59% in vitro inhibition of ACE and renin activities, respectively in comparison to the membrane fractions with 46-78 and 41-66% values. The ACE-inhibitory IC50 values were 0.22-0.79 mg/mL when compared to 0.41-0.96 mg/mL for renin, which suggests that the peptides have higher potency against ACE. The SnPH and membrane fractions inhibited ACE activity through mainly a non-competitive mechanism whereas renin inhibition was of the mixed-type. The results suggest that the snail peptides bind mainly to ACE non-active sites but could bind to both the active and non-active sites of renin.
Keywords: Snail; Protein hydrolysate; Angiotensin converting enzyme; Renin; Enzyme inhibition kinetics; Membrane ultrafiltration; IC50 values
Introduction
Hypertension is defined as sustained increases in systolic blood pressure (SBP) of 140 mmHg or greater and/or diastolic blood pressure (DBP) >90 mmHg. Hypertension is classified into two types: primary or essential hypertension (no underlying cause) and secondary hypertension (hypertension due to other disorders like kidney disease, vascular or endocrine disorders) [1,2]. Hypertension in human beings is controlled mainly through the renin-angiotensin system (RAS), which has become a critical physiological target for the development of anti-hypertensive and other cardio-protective agents. The RAS plays vital roles in the progression of human cardiovascular and chronic kidney diseases [3,4]. The RAS pathway is associated with series of enzyme-catalyzed reactions that produce compounds capable of regulating human blood pressure. Renin and angiotensin I-converting enzyme (ACE) are the two principal enzymes involved in RAS control. The key reactions in the RAS pathway involve renin (an aspartyl protease), which catalyzes the first and rate-limiting step by converting angiotensinogen to angiotensin I (AT-I), a decapeptide [5]. This is followed by ACE (peptidyldipeptidase A) that converts AT-I to a potent vasoconstrictor octapeptide called angiotensin II (AT-II). ACE also catalyzes bradykinin (a vasodilator) degradation and inactivation, which contributes to increased vasoconstriction [5]. Several ACE inhibitors such as captopril, enalapril, lisinopril, quinapril, ramipril, perindopril, and benazepril in addition to the only commercially approved renin inhibitor (aliskiren) as well as angiotensin receptor blockers (ARB) have found relevant clinical applications as drugs in hypertension treatment. However, prolonged usage of these drugs has been reported to be associated with undesirable side effects in some patients, which undermines compliance with physician-prescribed dosage [6,7].
Therefore, it has become increasingly necessary to discover safer and cheaper therapeutic compounds from natural food sources for lowering human blood pressure during hypertension. Moreover, it has been postulated that the direct renin activity inhibition could provide a better control of elevated blood pressure than ACE only inhibition (mono-therapy). This is because renin inhibition will lead to a reduction in AT-I (ACE substrate) level, which would otherwise be converted to AT-II in some organs via an ACEindependent pathway catalyzed by the passive action of chymase [8]. It must be noted however, that renin inhibition does not prevent the progression of ACE-catalyzed bradykinin degradation. Thus even in the presence of renin inhibitors, ACE-catalyzed bradykinin degradation may continue to cause blood vessel stiffening, which could ultimately contribute to elevated blood pressure [9]. Therefore, there is growing and urgent need to develop antihypertensive agents that will exert multi-enzyme inhibitory effects, especially with the capacity to simultaneously inhibit ACE and renin activities. Such multi-enzyme inhibition could enable more effective controls of elevated blood pressure, which will contribute to a reduced risk of hypertension development. Previous works have reported food protein hydrolysates with in vitro inhibition of both ACE and renin activities [10-12]. The in vitro renin and/or ACE inhibitions were also shown to be associated with lowering of elevated blood pressure in spontaneously hypertensive rats [12-16] and in hypertensive humans [17,18].
The health promoting benefits of food protein-derived peptides have remained a subject of interest to many food and nutrition scientists who are continually exploring new and underutilized food protein sources. This is especially true for the non-conventional protein sources that can be used for isolation of bioactive compounds to serve as therapeutic agents in hypertension.The African giant land snail (Archachatina marginata) is a soft-bodied type of mollusk that is composed basically of a head with a flattened foot inside a protective calcified shell. The African giant land snail is a non-conventional wildlife dietary meat with high protein and iron contents that is particularly relished by the African consumers as a delicacy meal [19]. Proximate composition studies of the most popular species of edible African giant land snails have shown them to have 17-21% crude protein contents [20,21]. The snail protein compares well with other conventional livestock, meats like mutton, duck and chicken, which have crude protein values of 16.9%, 18.6% and 20.5%, respectively. In addition, snail proteins possess an excellent amino acid profile that could enhance isolation of peptides with the required amino acids for antihypertensive activity. Snails are cheap to rear both at subsistent and commercial levels with high returns on low input and could serve as a valuable source of raw materials for nutritional products [21]. However, to date, there is no information on the potential enzymatic release of bioactive peptides from snail muscle proteins. Therefore, the objective of this work was to determine the in vitro ACE and renin-inhibitory activities as well as the enzyme inhibition kinetics of African giant land snail protein-derived peptides.
Materials and Methods
Materials
Dried (<12% moisture content) ground African giant land snail protein meal (SnPM) was obtained from the Department of Wild Life and Range Management, University of Agriculture, Makurdi, Nigeria. Renin inhibitor screening assay kit was purchased from Cayman Chemicals (Ann Arbor, MI). Rabbit lung ACE, N-[3-(2-Furyl) acryloyl]-L-phenylalanyl-glycyl-glycine (FAPGG), pepsin (porcine gastric mucosa), and pancreatin (porcine pancrease) were purchased from Sigma-Aldrich (St. Louis, MO). Other analytical-grade reagents and ultrafiltration membranes (1, 3, 5, and 10 kDa molecular weight cut-off) were obtained from Fisher Scientific (Oakville, ON, Canada).
Preparation of snail protein hydrolysate and ultrafiltration membrane fractions
Prior to enzymatic hydrolysis, the ground SnPM was first defatted using acetone (1:10, w:v); the mixture was then stirred in the fume hood for 3 h and decanted followed by a second and a third consecutive extractions of the residue. The defatted SnPM was air-dried overnight in the fume hood and used for further studies. The defatted SnPM was enzymatically hydrolyzed according to a previous method [22] as follows. Briefly, a 5% (w/v, protein basis) defatted SnPM slurry was heated to 37 °C and adjusted to pH 2.0 using 2 M HCl. Protein hydrolysis was initiated by pepsin (4% w/v, protein basis) addition and the mixture stirred for 2 h. After peptic hydrolysis, the reaction mixture was adjusted to pH 7.5 with 2 M NaOH, pancreatin (4% w/v, protein basis) was then added and the mixture incubated at 37 °C for 4 h. The enzymatic reaction was terminated by adjusting the mixture to pH 4.0 with 2 M HCl followed by heating to 95 °C for 15 min to ensure a complete denaturation of residual enzymes. The mixture was centrifuged (7000g at 4 °C) for 30 min and the resulting supernatant was labeled snail protein hydrolysate (SnPH). The SnPH was sequentially passed through ultrafiltration membranes with molecular weight cut-off (MWCO) of 1, 3, 5, and 10 kDa in an Amicon stirred ultrafiltration cell. The retentate from 1 kDa was passed through the 3 kDa membrane whose retentate was passed through the 5 kDa with the retentate passed through the 10 kDa membrane; the permeate from each membrane was collected to obtain <1, 1-3, 3-5, 5-10 kDa peptide fractions, respectively. Permeates and the 10 kDa membrane retentate (peptides >10 kDa) were collected, lyophilized and stored at -20 °C. The above digestion and ultrafiltration membrane fractionation were performed in triplicate and the freeze-dried products combined and used for in vitro screening of renin and ACE-inhibitory properties.
Determination of protein content and yield of snail peptides
The percent protein contents of the SnPM, SnPH and its ultrafiltration membrane fractions were determined by modified Lowry method [23] while the yield for the SnPH and its membrane peptide fractions were determined according previously described procedures [22]. Briefly, SnPH yield was determined as the ratio of peptide weight of lyophilized SnPH to the protein weight of unhydrolyzed SnPM. Similarly, the percent yields of the ultrafiltration membrane fractions were calculated as the ratio of the respective weights of the lyophilized peptide permeates to the SnPH peptide weight.
ACE inhibition assay
The ability of snail peptide samples to inhibit ACE in vitro activity was measured according to a previously described spectrophotometric method that uses FAPGG as a substrate [24]. Briefly, 1 mL of 0.5 mM FAPGG (dissolved in 50 mM Tris-HCl buffer containing 0.3 M NaCl, pH 7.5) was mixed with 20 μL of ACE (1 U/mL, final activity of 20 mU) and 200 μL of sample dissolved in the same buffer as the FAPGG. The rate of decrease in absorbance at 345 nm was recorded for 2 min at room temperature. In the blank experiment, the buffer was used instead of peptide solution. ACE activity was expressed as rate of reaction (ΔA/min) and inhibitory activity was calculated as:
ACE inhibition (%) = [1 –ΔA.min-1 (sample)/ ΔA.min-1 (blank)] × 100
Where ΔA.min-1 (sample) and ΔA.min-1 (blank) are ACE activity in the presence and absence of inhibitory peptides, respectively.
Renin inhibition assay
In vitro assay of human recombinant renin activity was conducted using the Renin Inhibitor Screening Assay Kit according to a previously described method [25]. Briefly, snail peptide samples were diluted in Tris-HCl buffer (50 mM, pH 8.0, containing 100 mM NaCl), and pre-warmed to 37 °C. Before the reaction was started, (1) 20 μL substrate, 160 μL assay buffer, and 10 μL Double Distilled Water (DDW) were added to the background wells; (2) 20 μL substrate, 150 μL assay buffer, and 10 μL DDW were added to the control wells; and (3) 20 μL substrate, 150 μL assay buffer, and 10 μL sample were added to the inhibitor (sample) wells. The reaction was initiated by adding 10 μL renin to the control and sample wells. The microplate was shaken for 10 s for proper mixing and incubated at 37 °C for 15 min; fluorescence intensity (FI) was then recorded at excitation wavelength of 340 nm and emission wavelength of 490 nm using a fluorometric microplate reader (Spectra MAX Gemini, Molecular Devices, Sunnyvale, CA). The percentage renin inhibition was calculated as follows:
Renin inhibition (%) = [1 –ΔFIU.min-1 (sample)/ ΔFIU.min-1 (blank)] x 100
Where ΔFIU.min-1 (sample) and ΔFIU.min-1 (blank) are renin activity in the presence and absence of inhibitory peptides, respectively.
Kinetics studies of ACE and renin inhibition
The concentration of snail peptide that inhibited ACE activity by 50% (IC50) was calculated by non-linear regression from a plot of percentage ACE inhibition versus four peptide concentrations (0.125, 0.25, 0.5, and 1.0 mg/mL). The kinetics of ACE inhibition was studied with 0.0625, 0.125, 0.25 and 0.5 mM substrate (FAPGG) concentrations. The mode of ACE inhibition was determined from the Lineweaver-Burk plots while kinetic parameters (Vmax and Km) were estimated from non-linear regression fit of the data to the Michaelis-Menten equation using GraphPad Prism version 5.0 (GraphPad Software, San Diego, CA, USA). Inhibition constant (Ki) was calculated as the x-axis intercept from a plot of the slope of the Lineweaver-Burk lines against sample concentration while the catalytic efficiency (CE) was calculated fromVmaxx/Km ratio. Similarly for renin kinetics, the IC50 was calculated using the non-linear regression from a plot of percentage renin inhibition versus snail peptide concentrations (0.125, 0.25, 0.5, and 1.0 mg/mL). The renin inhibition kinetics was conducted using five (0.625, 1.25, 2.5, 5 and 10 μM) substrate concentrations in the absence and presence of samples while kinetic parameters were calculated as described above for ACE.
Statistical analysis
All assays were conducted in triplicate and analyzed by one-way analysis of variance (ANOVA). The means were compared using Duncan’s multiple range test and significant differences accepted at p<0.05.
Results and Discussion
Protein content and yield of SnPH and its ultrafiltration membrane peptide fractions
Enzymatic hydrolysis of the African giant land SnPM was carried out to mimic human gastrointestinal tract (GIT) digestion using a combination of pepsin and pancreatin proteases to produce SnPH. Protein contents were determined to be 18, 81, 68, 71, 73, 75 and 76% for SnPM, SnPH, <1 kDa, 1-3 kDa, 3-5 kDa, 5-10 kDa and >10 kDa samples, respectively (Table 1). The results indicate that enzymatic hydrolysis and the isolation methods enhanced the separation of peptides from non-protein materials, hence the significantly higher protein contents of the hydrolyzed products. The results confirm susceptibility of the snail proteins to enzyme-induced release of peptides, which could enhance value-added utilization of the snail meat. The comparatively lower 68% protein content of the initial peptide permeate (<1 kDa peptides) may be attributed to the presence of impurities, especially salt (NaCl) since the membrane was used first. Most of the salts are formed during the protein hydrolysis steps when the addition of NaOH is necessary to maintain reaction pH. The results are similar to a previous report on hemp seed peptides, which obtained a lower protein content for the <1 kDa peptides when compared to >1 kDa peptide fractions [22].
![]()
Sample
Protein content (%)
Yield (%)*
SnPM
18.0 ± 0.21
N/A
SnPH
81.0 ± 0.25
76.0 ± 0.59
<1 kDa
68.0 ± 1.20
40.0 ± 0.31
1-3 kDa
71.0 ± 0.33
25.0 ± 0.60
3-5 kDa
73.0 ± 0.42
18.0 ± 0.38
5-10 kDa
75.0 ± 0.32
10.0 ± 0.87
>10 kDa
76.0 ± 0.34
7.0 ± 0.25
*SnPH yield was based on SnPM while the peptide fractions yields were based on SnPH
Table 1: Protein content and yield of snail protein meal (SnPM), hydrolysate (SnPH) and membrane ultrafiltration peptide fractions.
Table 1 also shows the yield of SnPH and its ultrafiltration membrane peptide fractions. The percent yield is often used as a marker of the efficiency of the hydrolytic process towards peptide recovery. The yield also indicates protease effectiveness in peptide bond cleavage and release of a mixture of low molecular weight (<10 kDa) and high molecular weight (>10 kDa) peptide sequences. The yield could reveal the economic prospects of commercializing the protein hydrolysate as an ingredient for functional foods and nutraceuticals formulation. Higher peptide yields will be more beneficial for commercial processing and marketing of new products in contrast to low yielding samples that may not be economically viable. The SnPH peptide yield was 76%, which indicates about a quarter of the proteins in SnPH were not digested or some of the peptides were lost during processing. The high yield obtained in this work indicates increased hydrolytic efficiency associated with the use of two separate enzymes with different peptide bond cleavage specificities. It is possible that initial pepsin hydrolysis led to production of peptides that were more readily digested by pancreatin proteases hence the high peptide yield. The yield obtained in this work is less than the ~87% value reported for a hemp seed protein hydrolysate obtained from pepsin+pancreatin hydrolysis [22]. Thus, the snail proteins seem to be less digestible than hemp seed proteins, which reflects the higher protease susceptibility of the amorphous plant protein when compared to the more organized and fibrous animal protein. The 40% yield obtained for the <1 kDa fraction in this work is similar to the 42% reported for a <1 kDa hemp seed protein hydrolysate fraction [22]. Generally, the peptide yield decreased as the ultrafiltration membrane size increased, which is consistent with the high efficiency of the pepsin+pancreatin combination in reducing the snail proteins to small peptides. The results showed that 93% of the peptides in SnPH were of low molecular weight (<10 kDa) while 7% can be categorized as high molecular weight (>10 kDa) peptides. The dominance of low molecular weight peptides is highly desirable for bioactive peptides since the small sizes are believed to favour absorption from the GIT [5].
ACE-inhibitory properties of snail peptides
The practice of initial laboratory screening of samples for potential bioactivity is often used to determine potent samples that may be used for in vivo testing in appropriate animal models. The percentage ACE-inhibitory activities of snail peptides are shown in Figure 1, which indicates higher potency as peptide size increased from <1 kDa to >10 kDa. The results showed that membrane ultrafiltration fractionation led to improved ACE-inhibitory activities but only for the 5-10 and >10 kDa peptide fractions. The results also suggest that longer peptides had stronger interactions with ACE, which led to a more extensive reduction in catalytic activity when compared to shorter (<10 kDa) peptides. The results are in contrast to other reports that suggested peptides with <3 kDa sizes have better ACE-inhibitory activities than those with >3 kDa sizes. For example, the <1 kDa and 1-3 kDa ultrafiltration membrane peptide fractions obtained from hemp seed protein hydrolysate inhibited 40 and 36% ACE activity, respectively, in comparison to 29% for <3-5 kDa and 19% for 5-10 kDa [22]. Other reports from apricot (Prunus armeniaca L.) kernel protein hydrolysates [26] and ultrafiltration cowpea (Vigna unguiculata) protein hydrolysate peptide fractions [27] have also showed that smaller size peptides are superior inhibitors of ACE activity than the bigger size peptides. The ACE-inhibitory effects shown by the SnPH and its ultrafiltration membrane peptide fractions (46-78%) are similar to those of pancreatin-hydrolyzed fish muscle samples that had 69–77% [28]. However, the ACEinhibitory activities obtained in this work are lower than the percent inhibition reported for alcalase or chymotrypsin-derived Australian canola protein hydrolysates, which had 90 and 89% ACE-inhibitory activities, respectively [13]. Higher ACE-inhibitory activities of 85- 89% and 82% have also been reported for alcalase-derived rapeseed [12,29] and chicken skin [11] protein hydrolysates, respectively. These differences in ACE-inhibitory activity may be due to the use of different proteases as well as varied primary structure (amino acid sequence) of the native protein raw materials, which will lead to liberation of peptides that differ in amino acid composition and sequence.
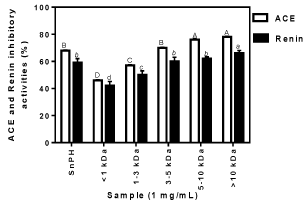
Figure 1: Percent in vitro Angiotensin Converting Enzyme (ACE) and renin
inhibitory activities of snail protein hydrolysate (SnPH) and peptide fractions.
The ultrafiltrated solution treated with pepsin and pancreatin, but without
SnPH had zero inhibitory effect against renin and ACE. Bars with different
letter shave significantly different (p<0.05) mean values.
Renin inhibitory activities of snail peptides
As shown in Figure 1, SnPH and all its membrane fractions exhibited moderate renin inhibitory activities which ranged from 42-66% and increased with increase in peptide size. The bigger size peptides (3-5, 5-10 and >10 kDa) possessed better renin-inhibitory properties (60, 62 and 66%, respectively) than the smaller size (<1 and 1-3 kDa) peptides, which exhibited lower inhibitory effects of 42 and 50%, respectively. Clearly, ultrafiltration membrane fractionation of SnPH (renin-inhibitory activity of 59%) resulted in significantly improved activity only when the peptide size was >10 kDa. These results are consistent with previous observations [30,31] that have indicated better renin-inhibitory effects for high MW than low MW compounds.
Inhibitory potency of snail peptides based on IC50 values
Figures 2A and B show the dose-response curve for ACE and renin inhibitions, respectively. The IC50values for the samples were then obtained by subjecting each line to a non-linear regression equation. As shown in Table 2, the IC50values of snail peptides for both ACE and renin inhibition correlated with their percent in vitro inhibitory effects. For instance the IC50values of ACE inhibition by snail peptides decreased with MW whereas the in vitro inhibitory effects were found to increase with MW. The lowest IC50values for ACE inhibition were exhibited by the bigger size peptides (3-5, 5-10 & >10 kDa), which were determined to be 0.22, 0.33 and 0.38 mg/ mL, respectively in comparison to the 0.79 mg/mL for <1 kDa and 0.76 mg/mL for 1-3 kDa peptides. The results indicate that smaller amounts of bigger-size peptides will be required to achieve maximal ACE-inhibitory effects when compared to the small size peptides. The ACE-inhibitory IC50value of SnPH (0.43 mg/mL) was also observed to be significantly (p<0.05) lower than those of the <1 kDa (0.79 mg/ mL) and 1-3 kDa (0.76 mg/mL) membrane fractions. The high yield and in vitro inhibitory effects associated with SnPH suggest that membrane ultrafiltration may not be required for commercialization purposes. Rather, the SnPH peptides could be a viable option for the industry due to lack of need for additional processing; the associated lower cost could enhance profitability and economic viability of the product. Nakajima et al. [28] previously reported an ACE-inhibitory IC50value of 0.79 mg/mL for pepsin+pancreatin hydrolyzed Atlantic salmon protein hydrolysate, which is similar to the IC50values of the <1 kDa and 1-3 kDa snail peptides (0.76-0.79 mg/mL). However, the snail peptide fractions have lower ACE-inhibitory IC50values (stronger potency) when compared to the in vitro hydrolyzed salmon myofibrillar and sarcoplasmic samples with IC50 values of 0.91 and 1.04 mg/mL, respectively [32]. The IC50values for renin inhibition by snail peptides also followed a similar trend as their percent inhibitory effects. As expected, the bigger size peptides (3-5, 5-10 and >10 kDa) had lower IC50values of 0.59, 0.47 and 0.41 mg/mL, respectively when compared to the higher values of 0.94 mg/mL for <1 kDa and 0.91 mg/mL for the 1-3 kDa peptide fractions. Overall, IC50values of renin inhibition by snail peptides were of higher magnitude (0.41-0.96 mg/ mL) than those of ACE inhibition (0.22-0.79 mg/mL), which confirm previous reports indicating ACE as a more readily inhibited enzyme than renin. There are no available renin-inhibitory IC50 values in literature for mollusks or their close relatives (marine peptides) that can be compared to current results for snail peptides. However, the renin-inhibitory IC50 values obtained in this work for snail peptides are lower than the 0.81, 1.89, and 2.52 mg/mL reported for a hemp seed protein hydrolysate, the <1 kDa fraction and the 1-3 kDa fraction, respectively.
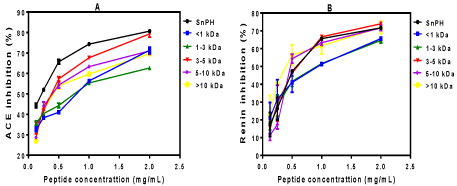
Figure 2: Dose-dependent changes in ACE (A) and Renin (B) inhibitions by snail protein hydrolysate (SnPH) and peptide fractions.
![]()
Sample
ACE
Renin
SnPH*
0.43± 0.01
0.58 ± 0.02
<1 kDa
0.79 ± 0.00
0.94 ± 0.03
1-3 kDa
0.76 ± 0.00
0.91 ± 0.01
3-5 kDa
0.38 ± 0.01
0.59 ± 0.02
5-10 kDa
0.33 ± 0.03
0.47 ± 0.01
>10 kDa
0.22 ± 0.12
0.41 ± 0.04
*Snail protein hydrolysate
Table 2: Peptide inhibitory concentration (mg/mL) that reduced enzyme activity by 50% (IC50).
Kinetics of ACE inhibition
The double reciprocal plots of ACE-catalyzed reactions in the absence and presence of snail peptides indicates a mostly noncompetitive type of inhibition as shown in Figure 3A-F for SnPH and its membrane peptide fractions. The results suggest that the snail peptides can bind to both the free enzyme as well as the enzymesubstrate complex. When bound to the free enzyme, the peptides will reduce substrate binding while binding to the enzyme substrate complex will reduce product formation. Since the peptides bind to both the free enzyme and the enzyme-substrate complex, the Km value will be very similar at different peptide concentrations as shown in Table 3. The decreased rate of catalysis is reflected in Catalytic Efficiency (CE) and maximum velocity (Vmaxx) decreases as shown also in Table 3, which confirms interaction of peptides with ACE to reduce product formation. The Km value (0.729 mM FAPGG) of the uninhibited ACE reaction in this study is higher than previously reported values of 0.306 mM for flaxseed protein hydrolysate [33], 0.664 mM for hemp seed protein hydrolysate [22] and 0.32 mM for Vernoniaamygdalina and Gongronemalatifolium leafy vegetable polyphenolic extracts [34]. The inhibition constant (Ki) is defined as a measure of the strength of peptide binding to ACE enzyme; therefore, low values indicate stronger binding affinity when compared to high values. The SnPH showed the highest binding affinity towards ACE with a Kiof 0.263 mg/mL followed by the bigger size peptides (3-5, 5-10 and >10 kDa) with a Ki range of 0.309-0.705 mg/mL. In contrast the <1 kDa and 1-3 kDa peptides had the least ACE-binding affinity with higher Kivalues of 0.725 and 0.830 mg/mL, respectively. The Ki values for SnPH and its membrane fractions obtained in this work are lower than the values (2.550-4.740 mg/mL) reported for hemp seed protein hydrolysate and its membrane fractions [22]. Several Lineweaver-Burk plot patterns for ACE inhibition by food proteinderived peptides have been previously reported. Non-competitive mode of ACE inhibition was reported for lentil protein hydrolysate [35] while pea protein [36] and chickpea protein [37] hydrolysates were both associated with uncompetitive inhibition of ACE enzyme. In contrast, the pepsin+pancreatin digested chicken skin peptides showed mixed-type pattern of ACE inhibition [38].

Figure 3: Lineweaver-Burk plots of angiotensin I-converting enzyme (ACE) inhibition by snail protein hydrolysate (SnPH) and its ultrafiltration peptide fractions.
![]()
Catalytic parameter
Peptide concentration (mg/mL)
Control
SnPH
<1 kDa
1-3 kDa
3-5 kDa
5-10 kDa
>10 kDa
0
0.3
0.6
0.4
0.8
0.5
1.0
0.5
1.0
0.8
1.5
0.8
1.5
Vmax
0.031
0.018
0.014
0.019
0.014
0.019
0.010
0.014
0.010
0.021
0.012
0.018
0.013
Km
0.729
0.753
0.831
0.770
0.765
0.738
0.575
0.745
0.700
0.873
0.727
0.627
0.615
CE
0.043
0.024
0.017
0.024
0.018
0.025
0.017
0.019
0.013
0.024
0.016
0.029
0.022
Ki
0.231
0.725
0.830
0.309
0.586
0.705
Km is Michaelis constants(mM); Vmax is maximum reaction velocity (ΔA/min); CE is the catalytic ef?ciency; Ki(mg/mL) is the enzyme-inhibitor dissociation constant
Table 3: Kinetics constants for ACE inhibition in the presence and absence of snail protein hydrolysate (SnPH) and membrane ultrafiltration peptide fractions.
Kinetics of renin inhibition
Unlike ACE kinetics studies in which SnPH and all its ultrafiltration membrane peptide fractions were evaluated for their respective mode of inhibiting ACE activity, the renin inhibition kinetics could only be determined with the most active samples (SnPH, 5-10 kDa and >10 kDa) as shown in Figure 4. Renin inhibition by the SnPH followed a mostly uncompetitive mode as shown by decreases in Vmaxx and Km values (Figure 4A and Table 4). The results suggest that SnPH peptides bind mostly to the renin-substrate complex to reduce rate of substrate conversion (lower Vmaxx) and increase substrate affinity (lower Km). Similar uncompetitive renin inhibition pattern has been reported for leaf polyphenols [34] and flaxseed peptides [33]. CE of the renin enzyme in the presence of different SnPH concentrations was significantly reduced from 12.06 (uninhibited reaction) to 6.323 and 4.742 for the low (0.8 mg/mL) and high (1.5 mg/mL) inhibitor concentrations, respectively (Table 4). Figure 4B shows that the 5-10 kDa peptides displayed non-competitive inhibition of renin activity, which suggests binding to non-active site only; hence similar Km values but reduced Vmaxx (Table 4). In contrast, the >10 kDa peptides exhibited mixed-type mode of renin inhibition, which was characterized by higher Kmvalue at 0.5 mg/mL but similar value at 1.0 mg/mL while reduced Vmaxx values were obtained at both peptide concentrations (Table 4). Therefore, the results suggest that the >10 kDa peptides could bind simultaneously to both the active site as well as the non-active sites to limit renin catalytic rate. The mixed-type mechanisms of renin inhibition displayed by the >10 kDa peptides is similar to those reported for hemp seed protein hydrolysate peptides [22]. The decreases in CE in the presence of snail peptides confirm interactions with renin protein to limit proteolytic efficiency.
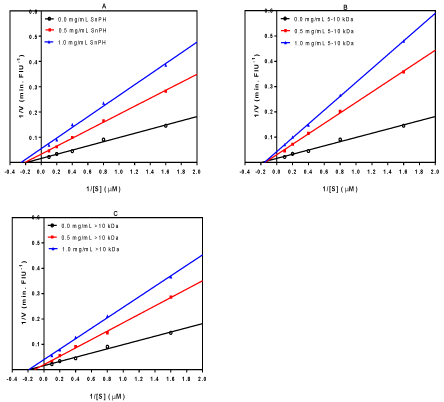
Figure 4: Lineweaver-Burk plots of renin inhibition of by snail protein hydrolysate (SnPH) and its ultrafiltration peptide fractions.
![]()
Catalytic parameter
Peptide concentration (mg/mL)
Control
SnPH
5-10 kDa
>10 kDa
0
0.8
1.5
0.5
1.0
0.5
1.0
Vmax
62.84
30.34
18.31
32.92
23.44
50.63
25.24
Km
5.21
4.80
3.61
6.81
6.41
8.39
5.22
CE
12.06
6.32
4.74
4.84
3.66
6.04
4.84
Ki
1.30
1.04
1.51
Km is Michaelis constants(mM); Vmax is maximum reaction velocity (fluorescenceintensity units/min); CE is the catalytic ef?ciency; Ki(mg/mL) is the enzyme-inhibitor dissociation constant
Table 4: Kinetics constants for renin inhibition in the presence and absence of snail protein hydrolysate (SnPH) and membrane ultrafiltrationpeptide fractions.
Conclusion
The study confirms the possibility of producing low molecular weight peptides from enzymatic hydrolysis of snail muscle proteins. The high peptide yield makes the process compatible with potential commercialization since it could enhance profitability. The results indicate that snail protein-derived peptides can reduce in vitro catalytic activities of ACE and renin, the main causative agents of hypertension. Inhibition of ACE activity was higher than that of renin activity, which confirms that renin protein conformation presents a more difficult conformation for inhibitors to overcome. The ease of ACE inhibition was also reflected in the fact that all the peptides acted mostly through non-competitive enzyme inhibition, i.e., binding to the non-active site. In contrast, the peptides inhibited the more difficult renin through binding to both the active and non-active sites. The ability to inhibit both renin and ACE activities suggest that snail peptides could potentially serve as therapeutic agents in the prevention and management of hypertension and associated morbidities. However, animal and human studies are required to confirm antihypertensive efficacy under physiological conditions.
Acknowledgement
This work was funded by the Natural Sciences and Engineering Research Council (NSERC) of Canada through a Discovery grant award to Dr. R.E. Aluko.
References
- Sultanpur C, Kumar S, Deepa K. Vaccines for hypertension disorder: Beneficial or detrimental. Singapore J Sci Res. 2011; 1: 13, 22.
- Popescu SM, Scrieciu M, Mercut V, Tuculina M, Dascalu I. Hypertensive patients and their management in dentistry. Int Scholarly Res Notices. 2013; 2013: Article ID 410740.
- DeMello WC, Re RN. Systemic versus local renin angiotensin systems: An overview. DeMello WC, Frohlich ED, eds. In: Renin Angiotensin System and Cardiovascular Disease. New York, Humana Press. 2009: 1-5.
- Daien V, Duny Y, Ribstein J, du Cailar G, Mimran A, Villain M, et al. Treatment of hypertension with renin-angiotensin system inhibitors and renal dysfunction: a systematic review and meta-analysis. See comment in PubMed Commons below Am J Hypertens. 2012; 25: 126-132.
- Aluko RE. Antihypertensive peptides from food proteins. See comment in PubMed Commons below Annu Rev Food Sci Technol. 2015; 6: 235-262.
- Antonios TF, MacGregor GA. Angiotensin converting enzyme inhibitors in hypertension: potential problems. See comment in PubMed Commons below J Hypertens Suppl. 1995; 13: S11-16.
- Abassi Z, Winaver J, Feuerstein GZ. The biochemical pharmacology of renin inhibitors: implications for translational medicine in hypertension, diabetic nephropathy and heart failure: expectations and reality. Biochem Pharmacol. 2009; 78: 933-940.
- Belova LA. Angiotensin II-generating enzymes. Biochemistry (Moscow) 2000; 65: 1589-1599.
- Segall L, Covic A, Goldsmith DJ. Direct renin inhibitors: the dawn of a new era, or just a variation on a theme? See comment in PubMed Commons below Nephrol Dial Transplant. 2007; 22: 2435-2439.
- Girgih AT, He R, Aluko RE. Kinetics and molecular docking studies of the inhibitions of angiotensin converting enzyme and renin activities by hemp seed (Cannabis sativa L.) peptides. See comment in PubMed Commons below J Agric Food Chem. 2014; 62: 4135-4144.
- Onuh JO, Girgih AT, Aluko RE, Aliani M. Inhibitions of renin and angiotensin converting enzyme activities by enzymatic chicken skin protein hydrolysates. Food Res Int. 2013; 53: 260-267.
- He R, Alashi A, Malomo SA, Girgih AT, Chao D, Ju X, et al. Antihypertensive and free radical scavenging properties of enzymatic rapeseed protein hydrolysates. See comment in PubMed Commons below Food Chem. 2013; 141: 153-159.
- Alashi AM, Blanchard CL, Mailer RJ, Agboola SO, Mawson AJ, He R, et al. Blood pressure lowering effects of Australian canola protein hydrolysates in spontaneously hypertensive rats. Food Res Int. 2014; 55: 281-287.
- He R, Malomo SA, Alashi A, Girgih AT, Ju X, Aluko RE. Purification and hypotensive activity of rapeseed protein-derived renin and angiotensin converting enzyme inhibitory peptides. J Funct Foods. 2013; 5: 781-789.
- Girgih AT, Alashi A, He R, Malomo S, Aluko RE. Preventive and treatment effects of a hemp seed (Cannabis sativa L.) meal protein hydrolysate against high blood pressure in spontaneously hypertensive rats. Eur J Nutr. 2014; 53: 1237-1246.
- Nwachukwu ID, Girgih AT, Malomo SA, Onuh JO, Aluko RE. Thermoase-derived flaxseed protein hydrolysates and membrane ultrafiltration peptide fractions have systolic blood pressure-lowering effects in spontaneously hypertensive rats. Int J Mol Sci. 2014; 15: 18131-18147.
- Li H, Prairie N, Udenigwe CC, Adebiyi AP, Tappia PS, Aukema HM, et al. Blood pressure lowering effect of a pea protein hydrolysate in hypertensive rats and humans. See comment in PubMed Commons below J Agric Food Chem. 2011; 59: 9854-9860.
- Seppo L, Jauhiainen T, Poussa T, Korpela R. A fermented milk high in bioactive peptides has a blood pressure-lowering effect in hypertensive subjects. See comment in PubMed Commons below Am J Clin Nutr. 2003; 77: 326-330.
- Babalola OO, Akinsoyinu AO. Proximate composition and mineral profile of snail meat from different breeds of land snail in Nigeria. Pakistan J Nutr. 2009; 8: 1842-1844.
- Ademolu K, Idowu A, Mafiana C, Osinowo O. Performance, proximate and mineral analyses of African giant land snail (Archachatina marginata) fed different nitrogen sources. Afr J Biotechnol. 2005; 3: 412-417.
- Eneji C, Ogogo A, Emmanuel-Ikpeme C, Okon O. Nutritional assessment of some Nigerian land and water snail species. Ethiopian J Environ Stud Manag. 2008; 1: 56-60.
- Girgih AT, Udenigwe CC, Li H, Adebiyi AP, Aluko RE. Kinetics of enzyme inhibition and antihypertensive effects of hemp seed (Cannabis sativa L.) protein hydrolysates. J Am Oil Chem Soc. 2011; 88: 1767-1774.
- Markwell MA, Haas SM, Bieber LL, Tolbert NE. A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples. See comment in PubMed Commons below Anal Biochem. 1978; 87: 206-210.
- Holmquist B, Bünning P, Riordan JF. A continuous spectrophotometric assay for angiotensin converting enzyme. See comment in PubMed Commons below Anal Biochem. 1979; 95: 540-548.
- Mundi S, Aluko RE. Inhibitory properties of kidney bean protein hydrolysate and its membrane fractions against renin, angiotensin converting enzyme, and free radicals. Austin J Nutr Food Sci. 2014; 2: 11.
- Zhu Z, Qiu N, Yi J. Production and characterization of angiotensin converting enzyme (ACE) inhibitory peptides from apricot (Prunus armeniaca L.) kernel protein hydrolysate. Eur Food Res Technol. 2010; 231: 13-19.
- Segura Campos MR, Chel Guerrero LA, Betancur Ancona DA. Angiotensin-I converting enzyme inhibitory and antioxidant activities of peptide fractions extracted by ultrafiltration of cowpea Vigna unguiculata hydrolysates. J Sci Food Agric. 2010; 90: 2512.
- Nakajima K, Yoshie-Stark Y, Ogushi M. Comparison of ACE inhibitory and DPPH radical scavenging activities of fish muscle hydrolysates. Food Chem. 2009; 114: 844-851.
- Mäkinen S, Johannson T, Vegarud Gerd E, Pihlava JM, Pihlanto A. Angiotensin I-converting enzyme inhibitory and antioxidant properties of rapeseed hydrolysates. J Funct Foods. 2012; 4: 575-583.
- Yuan L, Wu J, Aluko RE, Ye X. Kinetics of renin inhibition by sodium houttuyfonate analogs. See comment in PubMed Commons below Biosci Biotechnol Biochem. 2006; 70: 2275-2280.
- Yuan L, Wu J, Aluko RE. Size of the aliphatic chain of sodium houttuyfonate analogs determines their affinity for renin and angiotensin I-converting enzyme. Int J Biol Macromol 2007; 41: 274-280.
- Darewicz M, Borawska J, Vegarud GE, Minkiewicz P, Iwaniak A. Angiotensin I-converting enzyme (ACE) inhibitory activity and ACE inhibitory peptides of salmon (Salmo salar) protein hydrolysates obtained by human and porcine gastrointestinal enzymes. Int J Mol Sci. 2014; 15: 14077-14101.
- Udenigwe CC, Lin YS, Hou WC, Aluko RE. Kinetics of the inhibition of renin and angiotensin I-converting enzyme by flaxseed protein hydrolysate fractions. J Funct Foods. 2009; 1: 199-207.
- Ajibola CF, Eleyinmi AF, Aluko RE. Kinetics of the inhibition of renin and angiotensin i converting enzyme by polar and non-polar polyphenolic extracts of Vernonia amygdalina and Gongronema latifolium leaves. Plant Foods Hum Nutr. 2011; 66: 320-327.
- Barbana C, Boye JI. Angiotensin I-converting enzyme inhibitory properties of lentil protein hydrolysates: Determination of the kinetics of inhibition. Food Chem. 2011; 127: 94-101.
- Jakubczyk A, Baraniak B. Angiotensin I converting enzyme inhibitory peptides obtained after in vitro hydrolysis of pea (Pisum sativum var. Bajka) globulins. See comment in PubMed Commons below Biomed Res Int. 2014; 2014: 438459.
- Pedroche J, Yust MM, Girón-Calle J, Alaiz M, Millán F, Vioque J. Utilisation of chickpea protein isolates for production of peptides with angiotensin I-converting enzyme (ACE)-inhibitory activity. J Sci Food Agric. 2002; 82: 960-965.
- Onuh JO, Girgih AT, Malomo SA, Aluko RE, Aliani M.Kinetics of in vitro renin and angiotensin converting enzyme inhibition by chicken skin protein hydrolysates and their blood pressure lowering effects in spontaneously hypertensive rats. J Funct Foods. 2015; 14: 133-143.
Citation: Girgih AT, Nwachukwu ID, Iwar MI, Fagbemi TN and Aluko RE. Kinetics of Renin and Angiotensin Converting Enzyme Inhibition by African Giant Land Snail (Archachatina marginata) Protein Hydrolysate and Membrane Ultrafiltration Peptide Fractions. Austin J Nutri Food Sci. 2015; 3(4): 1071. ISSN: 2381-8980.