
Abstract
Hydrogen sulfide (H2S) with its characteristic odour has been known for centuries as a toxic gas for animals and human, especially at higher concentration. It has been also demonstrated that it can be phytototoxic resulting in disorders of the development but lower amounts of this gas may have beneficial effects on plants exposed to several abiotic stress factors such as salinity, drought, heat or heavy metal stress [1]. It seems that the activation of the antioxidant system triggerred by H2S is responsible of the tolerance in H2S-treated plants [2]. In this mini-review, we try to give an overview of the potentially positive and protective roles of H2S in plants under metallic stress.
Keywords: Hydrogen sulfide; Gasotransmitter; Heavy metal stress; Nitrooxidative stress; Tolerance
Abbreviations
AOA: Aminooxyacetic Acid; APX: Ascorbate Peroxidase; AsA: Ascorbic Acid; CAS: Cyanoalanine Synthase; CAT: Catalase; Chl: Chlorophyll; CO: Carbon Monoxide; CS: Cysteine Synthase; Cys: Cysteine; D-DES: D-Cysteine Desulfhydrase; DHAR: Dehydroascorbate Reductase; GPX: Glutathione Peroxidase; H2O2: Hydrogen Peroxide; L-DES: L-Cysteine Desulfhydrase; DES: Cysteine Desulfhydrase; GR: Glutathione Reductase; GSH: Glutathione (reduced); GST: Glutathione-S-Transferase; GYY4137: Morpholin-4-ium 4-methoxyphenyl (morpholino) Phosphinodithioate; H2S: Hydrogen Sulfide; HT: Hypotaurine; MDA: Malondialdehyde; MDHAR: Monodehydroascorbate Reductase; MT: Metallothionein; NaHS: Sodium hydrosulfide; NO: Nitric Oxide; PAG: Propargylglicine; PCs: Phytochelatins; POD: Peroxidase; Pro: Proline; ROS: Reactive Oxygen Species; RNS: Reactive Nitrogen Species; RSS: Reactive Sulphur Species; SOD: Superoxide dismutase; SNP: Sodium nitroprusside.
Introduction
Hydrogen sulfide (H2S), a colourless gas with a small molecular size and a typical odour reminding to rotten egg, has been known to be toxic for animal and human at higher levels. At the same time, it was discovered that the presence of this gas has effects on plants’ physiological processes, such as photosynthetic activity, development or stress tolerance [3,4]. Lower concentrations of exogenous H2S, mainly derived from sodium hydrosulfide (NaHS) as donor, might have beneficial influences on plant growth and development [1], while excess H2S can be deleterious, as it was demonstrated in Arabidopsis treated with 200-800 μM NaHS [5].
It is well-known that certain thermal fountains and the typically sulphur emitting vulcanic vents (solfataras) are natural sources of hydrogen sulfide but it can be produced swamps or sewers due to sulfate- or sulfur-reducing microorganisms [6]. Besides, several industrial technologies e.g. in oil refineries produce this gas as a byproduct causing serious toxicity problems [7].
H2S is the third gasotransmitter which was discovered in the 2000s and similar to Nitric Oxide (NO) and Carbon Monoxide (CO) it was firstly detected in animal and human [8]. Several studies confirmed that H2S arises endogenously mainly in cerebral tissues and has numerous functions e.g. in neuromodulation, cardiovascular or neurodegenarative diseases [3,4]. Up to now many researchers exhibited that this gaseous molecule also has a remarkable cue in plant physiological processes such as seed germination, morphogenesis of root, photosynthesis, senescence or stomatal closure [9,10]. Moreover, it might have a key role in plant tolerance against nitrooxidative stress trigerred by biotic e.g. pathogens, [11] or abiotic factors like drought, excess salinity or Heavy Metals (HMs) [12,13]. Since the emission and circulation of HMs in food chain are still great challenges globally, it is useful to overview the results about the changes of H2S level in plants exposed to HM stress and the potentially beneficial exogenous application of H2S, as well.
The Role of H2S in Plants at Physiological Conditions
In plants, cysteine Desulfhydrases (DES) are considered to be the key enzymes of H2S synthesis catalyzing the production of H2S from L- or D- Cysteine (L-Cys and D- Cys) [1]. L-Cys Desulfhydrases (L-DES, EC 4.4.1.28.) use L-Cys as substrate creating H2S, pyruvate and ammonia and are mainly located in the cytosol, nucleus and mitochondria, whilst D-Cys Desulfhydrases (D-DES, EC 4.4.1.15.) degrade D-Cys and can be found in mitochondria [2,4]. On the contrary, earlier summary by Guo et al. [3] reported that the main location of D-DES enzymes is the cytoplasm and L-DESs are localized in the chloroplasts and mitochondria. The activity of L-DES enzymes have been already characterized in tobacco, pumpkin, cucumber, Arabidopsis and Brassica napus [14,3]. Cytosolic D-DES activity was exhibited in pumpkin, cucumber, spinach, tobacco and Arabidopsis [3,4]. Another difference between L- and D-DES enzymes is that the activity of L-DESs can be inhibited using Aminooxyacetic Acid (AOA) while it has no influence on D-DESs [3]. Not only DES enzymes ara capable of producing H2S in plant cells, but chloroplastic sulfite reductase (SiR, EC 1.8.7.1), cyanoalanine synthase (CAS, EC 4.4.1.9) localized in cytoplasm and mitochondria and cytosolic Cys synthase (CS, EC 2.5.1.47) also produce hydrogen sulfide [2,4]. Recently, Corpas et al. [15] demonstrated the presence of H2S in plant peroxisomes, too.
Like the other gasotransmitters NO or CO, hydrogen sulfide has proved to be an essential participant of several physiological processes (e.g. germination, lateral root formation, stomatal closure) at „normal circumstances”. Li et al. [16] demonstrated that if Jatropha curcas seeds were pre-treated with Aminooxyacetic acid (AOA), an inhibitor of H2S biosynthesis, endogenous H2S content decreased and germination percentage was significantly lower than Control. The essential role of H2S in lateral root emergence was also supported by a study [17] which showed an interaction among H2S, NO and Ca2+ ions. Using the inhibitor of H2S synthesis, Propargylglicine (PAG) and the scavanger of H2S, Hypotaurine (HT) the number of lateral root primordia significantly reduced. It was also detected that the application of Sodium Nitroprusside (SNP) as NO donor enhanced the generation of endogenous H2S, especially in lateral root primordia.
Some reports have already presented the signaling function of H2S or the cooperation between H2S and NO in regulating the stomatal apertures [18,19,9]. Nonetheless, H2>S also displayed to be important in delayed flower opening [20] and postharvest senescence of fruits through mediating the level of ROS and activating antioxidant enzymes [21,6].
Besides, supra-optimal levels of H2S, mainly due to excess application of H2S donors, might be toxic for plants. The typical symptoms of H2S toxicity are lesions in leaves, retarded growth of plants (both shoot and root), decrement of chlorophyll content and consequently reduced photosynthetic efficiency [6,5]. Zhang et al. presented that toxic level of endogenous H2S, deriving from high NaHS concentration (200μM‹) may have inhibitory effect on primary root development via ROS overproduction.
Recent studies have confirmed the probable interaction among H2S and Reactive Oxygen Species (ROS), Reactive Nitrogen Species (RNS) [9,22] resulting in modulation of nitro-oxidative status of the plant cells. H2S itself is also regarded to be a member of Reactive Sulphur Species (RSS) [22] and beside Glutathione (GSH) and NO has an important role in posttranslational modification (namely persulfidation) of numerous proteins, as it was announced in case of Arabidopsis [23].
The Role of H2S in Plants under HM Stress
Many researchers ascertained that the increased emission of HMs to environment originating from different sources like geological or anthropogenic activities (e.g. mining, industrial, excess application of fertilizers and pesticides or contaminated irrigation water) might have several negative effects on plants. The main symptoms of HM stress and/or toxicity in plants are generally disturbances in photosynthesis, reduced germination rate, abnormal development of vegetative parts (root and/or shoot) [24]. It is well-known that not only the toxic metals like Cd, Pb, Hg or As can cause nitro-oxidative stress in the plant cells but the essential HMs (e.g. Fe, Ni, Zn, or Cu) at supra-optimal concentration also cause ROS/RNS overproduction and consequently affect the activity of the antioxidant defense system [25,26].
There are some results demonstrating that HM stress irrespectively of being essential or non-essential can alter positively or negatively H2S production in the plant organ investigated [27,28].
Beneficial Effects of Exogenous H2S Treatment under HM Stress
Numerous studies have supported that the application of H2S donors, like the most frequently used sodium hydrosulfide (NaHS) or the slow-releasing GYY4137 (morpholin-4-ium 4-methoxyphenyl(morpholino) phosphinodithioate) at a certain concentration may have significantly positive effects on seed germination [16,29, 14,30,31], lateral root formation [17], stomatal closure [18], as well as stress tolerance against salinity [32,33,12], heavy metals [34,35], drought [12,36], heat [37,38] and chilling [39,12] (Figure 1).
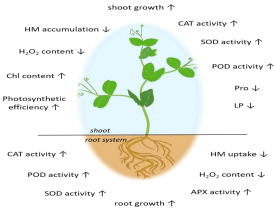
Figure 1: The effects of exogenous H2S in HM stressed plants.
The results of the reports reviewed show that in plants HM stress is usually accompanied by nitro-oxidative burst which is realized in Lipid Peroxidation (LP), the accumulation of ROS/RNS and the activation of the antioxidant system, but generally it becomes wellbalanced due to pre- or co-application of H2S, both in root and shoot (Table 1).
NaHS pre- or co-treatment at various concentrations seemed to be advantageous for the plant in point of HM uptake by root and accumulation in shoot. Exogenous H2S alleviated the uptake/ accumulation of essential HM Zn in black nightshade [40, Table 1] or toxic HM Cd and Pb in rapeseed [34,35,41,28], Cr in barley [42] and Arabidopsis [43].
Since, aluminum (Al) is not a HM considering its density (2.70gcm-3), but because of its toxic effects it is often regarded as HM [44], therefore we also take it into account. Exogenous H2S also mitigated the uptake/accumulation of Al in rapeseed and barley [45,46, 58,59] (Table 1).
Moreover, the application of H2S resulted in the increment Chlorophyll (Chl) and carotenoid content in the shoots (Table 1), as well, so it seems to have protective influence on HM stressed plants. The schematic model summarizes the main beneficial effects of exogenous H2S treatment.
References
- Li ZG. Hydrogen sulfide: a multifunctional gaseous molecule in plants. Russian Journal of Plant Physiology. 2013; 60: 733-740.
- He H, Li Y, He LF. The central role of hydrogen sulfide in plant responses to toxic metal stress. Ecotoxicology and Environmental Safety. 2018; 157: 403-408.
- Guo H, Xiao T, Zhou H, Xie Y, Shen W. Hydrogen sulfide: a versatile regulator of environmental stress in plants. Acta Physiologiae Plantarum. 2016; 38: 16.
- Corpas FJ, González-Gordo S, Cañas A, Palma JM. Nitric oxide and hydrogen sulfide in plants: which comes first?. Journal of Experimental Botany. 2019b.
- Zhang P, Luo Q, Wang R, Xu J. Hydrogen sulfide toxicity inhibits primary root growth through the ROS-NO pathway. Scientific Reports. 2017; 7: 868.
- Quirós-Sauceda AE, Velderrain-Rodríguez GR, Ovando-Martínez M, Goñi MG, González-Aguilar GA, Ayala-Zavala JF. Hydrogen Sulfide. In: Post harvest Management Approaches for Maintaining Quality of Fresh Produce (37-50). Springer. Cham. 2016; 39-50.
- Hendrickson RG, Chang A, Hamilton RJ. Co-worker fatalities from hydrogen sulfide. American Journal of Industrial Medicine. 2004; 45: 346-350.
- Wang R. Hydrogen sulfide: the third gasotransmitter in biology and medicine. Antioxidants & Redox Signaling. 2010; 12: 1061-1064.
- Hancock JT, Whiteman M. Hydrogen sulfide and cell signaling: team player or referee?. Plant Physiology and Biochemistry. 2014; 78: 37-42.
- Jin Z, Pei Y. Physiological implications of hydrogen sulfide in plants: pleasant exploration behind its unpleasant odour. Oxidative Medicine and Cellular Longevity. 2015.
- Li ZG, Min X, Zhou ZH. Hydrogen sulfide: a signal molecule in plant crossadaptation. Frontiers in Plant Science. 2016; 7: 1621.
- Shi H, Ye T, Han N, Bian H, Liu X, Chan Z. Hydrogen sulfide regulates abiotic stress tolerance and biotic stress resistance in Arabidopsis. Journal of Integrative Plant Biology. 2015; 57: 628-640.
- Hancock JT. Hydrogen sulfide and environmental stresses. Environmental and Experimental Botany. 2019; 161:50-56.
- Baudouin E, Poilevey A, Hewage NI, Cochet F, Puyaubert J, Bailly C. The significance of hydrogen sulfide for Arabidopsis seed germination. Frontiers in Plant Science. 2016; 7: 930.
- Corpas FJ, Barroso JB, González-Gordo S, Muñoz-Vargas MA, Palma JM. Hydrogen sulfide: A novel component in Arabidopsis peroxisomes which triggers catalase inhibition. Journal of Integrative Plant Biology Issue. 2019a; 61: 871-883.
- Li ZG, Gong M, Liu P. Hydrogen sulfide is a mediator in H2O2-induced seed germination in Jatropha curcas. Acta Physiologiae Plantarum. 2012a; 34: 2207-2213.
- Li YJ, Chen J, Xian M, Zhou LG, Han FX, Gan LJ, et al. In site bioimaging of hydrogen sulfide uncovers its pivotal role in regulating nitric oxide-induced lateral root formation. PLoS One. 2014; 9: 90340.
- Liu J, Hou L, Liu G, Liu X, Wang X. Hydrogen sulfide induced by nitric oxide mediates ethylene-induced stomatal closure of Arabidopsis thaliana. Chinese Science Bulletin. 2011; 56: 3547-3553.
- Hou Z, Wang L, Liu J, Hou L, Liu X. Hydrogen sulfide regulates ethyleneinduced stomatal closure in Arabidopsis thaliana. Journal of Integrative Plant Biology. 2013; 55: 277-289.
- Zhang H, Hu SL, Zhang ZJ, Hu LY, Jiang CX, Wei ZJ, et al. Hydrogen sulfide acts as a regulator of flower senescence in plants. Postharvest Biology and Technology. 2011; 60: 251-257.
- Ni ZJ, Hu KD, Song CB, Ma RH, Li ZR, Zheng JL, et al. Hydrogen sulfide alleviates postharvest senescence of grape by modulating the antioxidant defenses. Oxidative Medicine and Cellular Longevity. 2016.
- Corpas FJ. Nitric Oxide and Hydrogen Sulfide in Higher Plants under Physiological and Stress Conditions. Antioxidants. 2019; 8: 457.
- Aroca A, Gotor C, Romero LC. Hydrogen sulfide signaling in plants: Emerging roles of protein persulfidation. Frontiers in Plant Science. 2018; 9: 1369.
- Emamverdian A, Ding Y, Mokhberdoran F, Xie Y. Heavy metal stress and some mechanisms of plant defense response. The Scientific World Journal. 2015.
- Janas KM, Zielinska-Tomaszewska J, Rybaczek D, Maszewski J, Posmyk MM, Amarowicz R, et al. The impact of copper ions on growth, lipid peroxidation, and phenolic compound accumulation and localization in lentil (Lens culinaris Medic.) seedlings. Journal of Plant Physiology. 2010; 167: 270-276.
- Anjum NA, Umar S, Iqbal M, Khan NA. Cadmium causes oxidative stress in mung bean by affecting the antioxidant enzyme system and ascorbateglutathione cycle metabolism. Russian Journal of Plant Physiology. 2011; 58: 92-99.
- Chen Z, Yang B, Hao Z, Zhu J, Zhang Y, Xu T. Exogenous hydrogen sulfide ameliorates seed germination and seedling growth of cauliflower under lead stress and its antioxidant role. Journal of Plant Growth Regulation. 2018; 37: 5-15.
- Yu Y, Zhou X, Zhu Z, Zhou K. Sodium Hydrosulfide Mitigates Cadmium Toxicity by Promoting Cadmium Retention and Inhibiting Its Translocation from Roots to Shoots in Brassica napus. Journal of Agricultural and Food Chemistry. 2018; 67: 433-440.
- Dooley FD, Nair SP, Ward PD. Increased growth and germination success in plants following hydrogen sulfide administration. Plos One. 2013; 8: e62048.
- Carter JM, Brown EM, Grace JP, Salem AK, Irish EE, Bowden NB. Improved growth of pea, lettuce, and radish plants using the slow release of hydrogen sulfide from GYY-4137. PloS One. 2018; 13: e0208732.
- Chen Z, Yang B, Hao Z, Zhu J, Zhang Y, Xu T. Exogenous hydrogen sulfide ameliorates seed germination and seedling growth of cauliflower under lead stress and its antioxidant role. Journal of Plant Growth Regulation. 2018; 37: 5-15.
- Christou A, Manganaris GA, Papadopoulos I, Fotopoulos V. Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defence pathways. Journal of Experimental Botany. 2013; 64:1953-1966.
- Chen J, Wang WH, Wu FH, He EM, Liu X, Shangguan ZP, et al. Hydrogen sulfide enhances salt tolerance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Scientific Reports. 2015; 5: 12516.
- Ali B, Gill RA, Yang S, Gill MB, Ali S, Rafiq MT, et al. Hydrogen sulfide alleviates cadmium-induced morpho-physiological and ultrastructural changes in Brassica napus. Ecotoxicology and Environmental Safety. 2014a; 110: 197-207.
- Ali B, Mwamba TM, Gill RA, Yang C, Ali S, Daud MK, et al. Improvement of element uptake and antioxidative defense in Brassica napus under lead stress by application of hydrogen sulfide. Plant Growth Regulation. 2014b; 74: 261-273.
- Chen J, Shang YT, Wang WH, Chen XY, He EM, Zheng HL, et al. Hydrogen sulfide-mediated polyamines and sugar changes are involved in hydrogen sulfide-induced drought tolerance in Spinacia oleracea seedlings. Frontiers in Plant Science. 2016; 7: 1173.
- Li ZG, Long WB, Yang SZ, Wang YC, Tang JH, Wen L, et al. Endogenous hydrogen sulfide regulated by calcium is involved in thermotolerance in tobacco Nicotiana tabacum L. suspension cell cultures. Acta Physiologiae Plantarum. 2015a; 37: 219.
- Li ZG, Xie LR, Li XJ. Hydrogen sulfide acts as a downstream signal molecule in salicylic acid-induced heat tolerance in maize (Zea mays L.) seedlings. Journal of Plant Physiology. 2015b; 177: 121-127.
- Fu P, Wang W, Hou L, Liu X. Hydrogen sulfide is involved in the chilling stress response in Vitis vinifera L. Acta Societatis Botanicorum Poloniae. 2013; 82: 295-302.
- Liu X, Chen J, Wang GH, Wang WH, Shen ZJ, Luo MR, et al. Hydrogen sulfide alleviates zinc toxicity by reducing zinc uptake and regulating genes expression of antioxidative enzymes and metallothioneins in roots of the cadmium/zinc hyperaccumulator Solanum nigrum L. Plant and Soil. 2015; 400: 177-192.
- Ali B, Song WJ, Hu WZ, Luo XN, Gill RA, Wang J, et al. Hydrogen sulfide alleviates lead-induced photosynthetic and ultrastructural changes in oilseed rape. Ecotoxicology and Environmental Safety. 2014c; 102: 25-33.
- Ali S, Farooq MA, Hussain S, Yasmeen T, Abbasi GH, Zhang G. Alleviation of chromium toxicity by hydrogen sulfide in barley. Environmental Toxicology and Chemistry. 2013; 32: 2234-2239.
- Fang H, Liu Z, Jin Z, Zhang L, Liu D, Pei Y. An emphasis of hydrogen sulfidecysteine cycle on enhancing the tolerance to chromium stress in Arabidopsis. Environmental Pollution. 2016; 213: 870-877.
- Huang W, Yang X, Yao S, LwinOo T, He H, Wang A, et al. Reactive oxygen species burst induced by aluminum stress triggers mitochondria-dependent programmed cell death in peanut root tip cells. Plant Physiology and Biochemistry. 2014; 82: 76-84.
- Dawood M, Cao F, Jahangir MM, Zhang G, Wu F. Alleviation of aluminum toxicity by hydrogen sulfide is related to elevated ATPase, and suppressed aluminum uptake and oxidative stress in barley. Journal of Hazardous Materials. 2012; 209: 121-128.
- Ali B, Qian P, Sun R, Farooq MA, Gill RA, Wang J, et al. Hydrogen sulfide alleviates the aluminum-induced changes in Brassica napus as revealed by physiochemical and ultrastructural study of plant. Environmental Science and Pollution Research. 2015; 22: 3068-3081.
- Shan C, Dai H, Sun Y. Hydrogen sulfide protects wheat seedlings against copper stress by regulating the ascorbate and glutathione metabolism in leaves. Aust J Crop Sci. 2012; 6: 248-254.
- Dai H, Xu Y, Zhao L, Shan C. Alleviation of copper toxicity on chloroplast antioxidant capacity and photosystem II photochemistry of wheat by hydrogen sulfide. Brazilian Journal of Botany. 2016; 39: 787-793.
- Kaya C, Ashraf M, Akram NA. Hydrogen sulfide regulates the levels of key metabolites and antioxidant defense system to counteract oxidative stress in pepper (Capsicum annuum L.) plants exposed to high zinc regime. Environmental Science and Pollution Research. 2018; 25: 12612-12618.
- Ahmad R, Ali S, Rizwan M, Dawood M, Farid M, Hussain A, et al. Hydrogen sulfide alleviates chromium stress on cauliflower by restricting its uptake and enhancing antioxidative system. Physiologia Plantarum. 2019.
- Fang H, Jing T, Liu Z, Zhang L, Jin Z, Pei Y. Hydrogen sulfide interacts with calcium signaling to enhance the chromium tolerance in Setaria italica. Cell Calcium. 2014; 56: 472-481.
- Zhang H, Hu LY, Li P, Hu KD, Jiang CX, Luo JP. Hydrogen sulfide alleviated chromium toxicity in wheat. Biologia Plantarum. 2010; 54: 743-747.
- Zhang L, Pei Y, Wang H, Jin Z, Liu Z, Qiao Z, et al. Hydrogen sulfide alleviates cadmium-induced cell death through restraining ROS accumulation in roots of Brassica rapa L. ssp. pekinensis. Oxidative Medicine and Cellular Longevity. Article ID: 804603. 2015.
- Kabala K, Zboinska M, Glowiak D, Reda M, Jakubowska D, Janicka M. Interaction between the signaling molecules hydrogen sulfide and hydrogen peroxide and their role in vacuolar H+ATPase regulation in cadmium-stressed cucumber roots. Physiologia Plantarum. 2019; 166: 688-704.
- Cui W, Chen H, Zhu K, Jin Q, Xie Y, Cui J, et al. Cadmium-induced hydrogen sulfide synthesis is involved in cadmium tolerance in Medicago sativa by reestablishment of reduced (Homo) glutathione and reactive oxygen species homeostases. PloS One. 2014; 9: pe109669.
- Yang L, Zeng J, Wang P, Zhu J. Sodium hydrosulfide alleviates cadmium toxicity by changing cadmium chemical forms and increasing the activities of antioxidant enzymes in salix. Environmental and Experimental Botany. 2018; 156: 161-169.
- Bharwana SA, Ali S, Farooq MA, Ali B, Iqbal N, Abbas F, et al. Hydrogen sulfide ameliorates lead-induced morphological, photosynthetic, oxidative damages and biochemical changes in cotton. Environmental Science and Pollution Research. 2014; 21: 717-731.
- Huo J, Huang D, Zhang J, Fang H, Wang B, Wang C, et al. Hydrogen sulfide: a gaseous molecule in postharvest freshness. Frontiers in Plant Science. 2018; 9: 1172.
- Xie Y, Lai D, Mao Y, Zhang W, Shen W, Guan R. Molecular cloning, characterization, and expression analysis of a novel gene encoding L-cysteine desulfhydrase from Brassica napus. Molecular Biotechnology. 2013; 54: 737-746.