
Special Article - Sugarcane Sustainable Production
Ann Agric Crop Sci. 2021; 6(2): 1073.
Vinasse Waste from Sugarcane-Based Bioethanol Production Plants Kills Moniliophthora Perniciosa, the Causative Agent of Cacao Witches' Broom Disease
Ferraz P1,2, Amorim-Rodrigues M1,2, Cassio F1,2 and Lucas C1,2*
1Institute of Science and Innovation for Bio-Sustainability (IB-S), University of Minho, Campus de Gualtar, Portugal
2Department of Biology, Centre of Molecular and Environmental Biology (CBMA), University of Minho, Campus de Gualtar, Portugal
*Corresponding author: Lucas C, Institute of Science and Innovation for Bio-Sustainability (IB-S), University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal
Received: February 18, 2021; Accepted: March 16, 2021; Published: March 23, 2021
Abstract
Bioethanol production based on sugarcane juice fermentation yields vinasse, a dark, dense liquid waste high in potassium. In Brazil, one of the world’s biggest bioethanol producer, vinasse is used for fertirrigation of the sugar-cane fields, replacing mineral soil amendments. Nevertheless, the production largely exceeds this application, prompting exacerbated usage, unduly long-term storage and even illegal dumping. Vinasse thus progressively became an environmental hazard, damaging soils and superficial and ground waters, adding a negative burden to a supposedly green-fuel industry. The solution resides in decreasing production yields and/or using it for yet other economically interesting applications. This work focuses on the latter. Vinasses from three bioethanol plants from Brazil were tested for their ability to contain the proliferation of Moniliophthora perniciosa, the aggressive filamentous fungus responsible for cacao Witches’ Broom Disease (WBD). This disease caused a severe economic fall-out in South American cacao producing regions, particularly seriously in Brazil. Immersing or spraying the mycelium with vinasse either kills the fungus or impedes its proliferation at varying time/dosage. Identically testing another genetically unrelated phytopathogen, showed this effect is not that of a generalized biocide/fungicide. Results suggest that vinasse could be used to contain/revert the prevalence of cacao’s WBD to manageable levels. Vinasse would thus shift from industrial waste with disposal-associated costs, to being a tool for the agronomic sustainability and revival of the South American regional cacao-dependent socio-economies.
Keywords: Vinasse; Bioethanol; Cacao; Witches’ broom disease; Moniliophthora perniciosa
Introduction
Brazil produces bioethanol mostly from yeast fermentation of sugarcane juice. The process involves the physical extraction of the juice from the cane which is then fermented by yeasts. After each fermentation cycle, the must is centrifuged, and the yeast biomass separated and recycled. The subsequent distillation process yields a dark liquid waste called vinasse [1]. Brazilian industry generates an average 12L of vinasse per litre of ethanol produced [2], meaning that each year the country generates around 300-400 billion litres of vinasse [1]. This residue has been used for more than 50 years for fertirrigation (fertilization + irrigation) of the sugarcane fields [1,3]. This appears to be a virtuous cycle, since vinasse is rich in potassium and other minerals [4], avoiding the need for chemical fertilization [5]. In the long run, vinasse is apparently beneficial for soil quality and sugarcane production yield [4,6-9]. But in reality, the amounts of vinasse produced per year are far bigger than the fields can absorb, leading to saturation. Additionally, the vinasse’s low pH (3.5-5.0) and the corrosive nature [10] are promoting undesirable changes of soil composition and physicochemical properties [3,11], and serious contamination of ground water [12]. These problems are exacerbated by inappropriate long-term storage and illicit discharges.
The most studied possibilities of vinasse utilization relate with its reuse in the energy equation of the ethanol plant [4,13]. Nevertheless, vinasse is rich in minerals and organic matter [1,14,15], around 100-130 g/L (COD) [14], and includes sugars, organic acids, ethanol and glycerol [1,15], which suggests vinasse should be fit for microbial growth. Studies proposing this possibility include the cultivation of economically promising filamentous fungi such as the edible Rhizopus oligosporus [15], or the biotechnology relevant Aspergilus oryzae [16,17] and Neurospora intermedia [18]. In the present work, we explored yet another possibility, that of using vinasse for controlling the proliferation of the cacao phytopathogen Moniliophthora perniciosa, while used for fertirrigation of the cacao fields, as it happens with sugarcane.
perniciosa is a filamentous fungus that causes a devastating disease in the cacao plant (Theobroma cacao) and fruit known as Witches’ Broom Disease [19]. This was responsible for the steep production fall-out in South and Central America countries, mainly in Brazil, where it posed a serious social-economic crisis in the cacao producing region. The Brazilian production fell 70% in a period of 10 years after the onset of the disease in 1989 [20,21]. This scenario had a huge impact on the regional and national economy, leading the country to shift from being the 2nd world producer to becoming a net importer of cacao beans [21,22].
perniciosa is a very aggressive fungus, virtually impossible to eradicate. Its exceptional virulence is intrinsically related to its ability to infect all the pant tissues in all the stages of the cacao plant life-cycle [20,23,24]. M. perniciosa is a hemibiotrophic fungus, which infectious cycle has two distinct phases: a biotrophic and a saprotrophic. After the initial infection, a series of cell death events occurs in the infected tissues causing these to become necrotic and to form a particular structure called dry broom [20]. Those necrotic or dead plant cells are then colonized by the fungus, and pink-coloured basidiocarps are produced. In alternating periods of drought and humidity, each basidiocarp can produce 2 to 3.5 million spores [25]. The spores are released mainly at night and are locally dispersed by water and over long distances by wind, and can remain latent in the soil or inside pruned plant branches for long periods of time [20,23].
Classical chemical fungicides, usually azole or copper-based compounds, are not effective against M. perniciosa [26]. Presently, cacao Witches’ Broom Disease has one single functional management technique, which consists in spraying the affected plantations with Tricovab®, a Trichoderma stromaticum live suspension (reviewed by [24]). This other fungus is very efficient in antagonizing M. perniciosa, but it has to be multiplied in the surface of rice grains, being supplied lyophilized, as a service on demand, by an agro-support governmental organization (www.ceplac.gov.br). This is therefore a very expensive and unsustainable solution, only possible since it is almost fully subsidized by the Brazilian government. Alternatives could consist in using other microbes, easier and cheaper to cultivate, as biofungicides (reviewed by [24]), or possibly implementing a sustainability-prone solution controlling the fungus with an inexpensive agro-friendly industrial waste that is produced in very high amounts, and that might cumulatively contribute with mineral amendments for the cacao plants.
Three different vinasses were tested against two strains of the fungus originating from Brazil via a credited international culture collection. The fungi did not grow on vinasse. Accordingly, both died upon immersing or spraying with two of the vinasse batches, although at varying time/dosage, and were inhibited from proliferating by the third batch. This killing effect of vinasse was not shared by another genetically distant phytopathogenic fungus, suggesting specificity. Results support the use of vinasse for fertirrigation of cacao plantations as a means to, in time, clean the resident fungus inoculum. If not eradicating WBD, vinasse should at least reduce it to manageable levels. On the other hand, this new utilization would contribute to the alleviation of the negative unsustainable burden vinasse associates to the bioethanol industry.
Materials and Methods
Moniliophthora perniciosa strains CBS 441.80 and 442.80, and Colletotrichum gloeosporioides CBS 100471 were purchased from CBS-KNAW Collections, The Netherlands. They were maintained at 4ºC on MEA (20g/L malt extract w/ 20g/L agar), and cultivated in the same medium at 30°C. Vinasse was obtained from Fermentec Lda (https://www.fermentec.com.br/), and originated from three different bioethanol producing plants in the State of São Paulo, Brazil: Usina Alta Mogiana (http://www.altamogiana.com.br/), Usina Batatais (http://www.usinabatatais.com.br/) and Usina da Pedra (https://www.pedraagroindustrial.com.br/). Vinasse was stored at 4ºC. Prior to utilization it was centrifuged for 30min at 13.000xg and 4°C to eliminate particles in suspension, and autoclaved (121ºC, 1 atm, 20min). Growth assays were performed inoculating (i) 20mL of vinasse (100% vinasse); (ii) 10mL of vinasse +10mL of upH20 (50% vinasse); or (iii) 5mL of vinasse +15mL of upH20 (25% vinasse) - in a glass tube (Ø 3cm; 13cm height) with a one-week ME-grown fungus agar plug of approximately 0.8x0.8 cm. Tubes were incubated at 30ºC and 200rpm orbital shaking. Cultures identically inoculated and incubated in liquid ME medium were used as control. Growth was assessed after 10 days, decanting the culture supernatant, and checking for mycelium development. The putative viability of any remaining fungal cells was assessed incubating this agar plug in MEA at 30ºC for 1 week. These assays were repeated in the same manner, using vinasse supplemented with 2% of malt extract as a supplementary nutrient source. Death along incubation time in vinasse was assessed using a fully-grown fungal culture previously grown in liquid ME medium in a glass tube (until the formation of a globular mycelia), which was then transferred to a glass tube containing 20mL of vinasse, under the same conditions described above. Each sample/incubation time corresponding to an independent tube identically inoculated from fully-grown fungal cultures. The incubation period varied between 1 and 10 days (T1 to T10), and at increasing time points, the mycelia was taken from the vinasse, gently wash with sterile water and its viability assayed in MEA at 30°C for 1 week. The death-inducing effect without immersing the cells in vinasse was performed by spraying vinasse on a MEA plate containing fully grown mycelia. The assay was performed for 10 days, applying a single spray at T0, or repeating it once each 24h. The plates were then incubated at 30°C for 3 days. All assays were performed at least in three independent replicates (n ≥3).
Results and Discussion
The vinasse used in this work originated from three bioethanol plants from the State of São Paulo in Brazil, all of which identically producing bioethanol from a sugarcane juice fermentation process. According to the literature, the composition and physical properties of vinasses do not differ significantly (e.g. [14,15]). The three vinasses used in this work have ±6 % solids in suspension (mostly ashes), pH ≈ 4.5, 0.115 ± 0.060 % (w/v) sugars and 0.9% (w/v) organic acids (information kindly supplied by Fermentec, Lda. (https://www.fermentec.com.br)), values that fall within published data.
Does vinasse inhibit mycelium development?
The pH of vinasse is adequate to support the development of M. perniciosa which optimal growth occurs at pH between 4.5 and 5.5 (our unpublished results), and the presence of carbon source (totaling ± 0.5% (w/v)) should allow some fungal multiplication. Therefore, vinasse was centrifuged, to remove the solids in suspension, and the remaining liquid fraction was sterilized and used to inoculate the two M. perniciosa strains. After 10 days (Figure 1a) there was no visible growth of either fungus. Microscopic inspection confirmed the absence of hyphae associated with the agar plugs used as inocula. To check whether there were still residual viable mycelia, the fungal plugs were taken from the vinasse (Figure 1a), re-inoculated in solid MEA medium, and incubated for a further 10 days at 30°C. As can be seen in Figure 1b, the plugs of the vinasses from Alta Mogiana and Batatais did not develop any growth, suggesting the fungus was killed. The fungal plug taken from the vinasse from Pedra developed mycelia, showing that it rather strongly inhibited mycelium development. These results show that vinasse from different plants, and possibly also different batches, has different effects over the Witches’ broom fungus, either killing it or just inhibiting its development.

Figure 1: Two M. perniciosa strains originating from contaminated cacao fruits and trees in Brazil were grown in unsupplemented vinasse originating from three Brazilian bioethanol plants. (A) Results were scored after a 10 days incubation at 30ºC, before and after decanting the vinasse showing the inoculum agar plug. Control growth in identical period and temperature in MEA showing the fungal biomass ball. (B) The plugs from (A) were inoculated in MEA solid medium. Only the plugs originating from the incubation in vinasse from Pedra showed mycelia growth.
Are the effects dependent on dose?
To verify if lower concentrations of vinasse were sufficient to exert these effects, the assays were repeated using the same 3 batches of vinasse (i) undiluted (100% vinasse); (ii) diluted 1:2 (50% vinasse) or (iii) diluted 1:4 (25% vinasse). Results are presented in Table 1. The vinasse from Pedra plant only inhibit the growth of either fungi undiluted, while the other two vinasses were effective against each of the fungal strains at different dilutions. The further assessment of the viability of residual mycelia was made as above. No fungal development was observed indicating that the concentrations of vinasse from Alta Mogiana and Batatais that were able to inhibit fungal growth also killed the fungus. This ability was therefore shown to be dose-dependent, suggesting it relies on a specific component in the vinasse, and depends on the relative amounts towards those of mycelium. This points to expect that differences in strength may possibly be found from batch to batch.
Is there a fungal growth-associated bias?
Considering that the carbon source present in vinasse might not be sufficient to allow the fungal to develop generously, and/or that the death of mycelia could occur at such different speed that the 10 days contact with the vinasse would not be enough to detect death of the mycelia, two different assays were performed. The first consisted in increasing the amount of carbon source to support survival and growth in vinasse by adding malt extract 2% (w/v) and repeating the incubation in the same conditions used in the previous assay. Results were identical to the previous ones (not shown), indicating that the growth inhibition observed when inoculating the fungal plugs in vinasse is not related with nutrient deprivation. The second approach aimed to establish how fast M. perniciosa was dying on vinasse. The assay consisted in having ME-grown mycelia and replacing ME with vinasse (Figure 2a). At increasing time points, the mycelia were taken from the vinasse and their viability was assayed in solid MEA medium as before. This was done using the vinasse originating from Alta Mogiana plant, for having proved able to kill the two fungal strains. Results showed that the complete death of the mycelia from the two fungal strains indeed occurred when the fungi were immersed in vinasse, but at different times. The strain CBS 441.80 lost full viability between day 6 and day 7 (Figure 2b), while the strain CBS 442.80 lost full viability faster, between day 2 and day 3 (Figure 2b). Vinasse has thus a different time/dosage effect on each of the two strains of M. perniciosa, which must be considered for the putative application in the field.
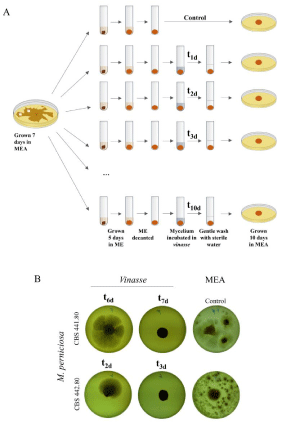
Figure 2: (A) Scheme of the procedure used to assay the effect of vinasse on the viability of the M. perniciosa along time. (B) Representative results of the turning point in the loss of viability which occurs at different time points for each fungal strain. The controls show the growth of the mycelium ball fragmented, which occurs whenever the inoculum is large.
Immersing or spraying?
To check whether the death-inducing effect could be obtained without the need to immerse the mycelium in vinasse, this was used to spray MEA fully grown mycelia. The spray was applied once every 24h for 10 days and the evolution of the mycelium was photographed (Figure 3). This time the three vinasses were used. Mycelium progressively shrunk, becoming darker coloured. Vinasses from the different plants had different time/dosage effect. The vinasse from Batatais killed the two strains faster than the one from Alta Mogiana, and Pedra’s presented the weaker inhibitory effect. Moreover, it was also possible to verify that each vinasse had a different time-effect on each of the M. perniciosa strains, being CBS 442.80 more sensitive since the mycelium disappeared between days 5 and 10 (except when using the vinasse from the Pedra plant). These results are consistent with the ones obtained by immersing mycelium in vinasse. Results further indicate that the spraying has to be repeated over time in order to observe the desired effect, corroborating the hypothesis that the continuity of the treatment could contribute to clean the field of hidden mycelium over time.
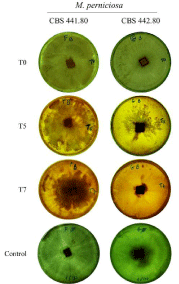
Figure 3: Chosen examples of the effect of spraying vinasse on the top of fully-grown mycelia on MEA once every 24h. Arrows indicate shrunk residual mycelium after 7 applications of vinasse.

Figure 4: C. gloeosporioides grown in unsupplemented vinasse originating from three different bioethanol plants from Brazil. Inoculum was done using a fully-grown mycelia agar plug as used for M. perniciosa. Results were scored after 10 days incubation at 30°C (1), and after decanting the remaining vinasse showing the fungal mycelium that was formed (2). Control growth was performed in MEA in identical period and temperature. C. gloeosporioides did not form a mycelium ball, instead it spread eventually filling the full liquid volume available.
Is the effect over M. perniciosa specific?
Finally, as an attempt to understand whether the ability of vinasse to kill filamentous fungi was specific for M. perniciosa and/or the correspondent taxa, the assays of fungal growth in vinasse for 10 days were repeated using another phytopathogenic fungus, Colletotrichum gloeosporioides [27]. This is one of the causative agents of olive anthracnose, a disease causing serious economic losses in the Mediterranean region [28] and it was chosen because it is a very resilient fungal species. Results (Figure 4) showed that C. gloeosporioides was able to grow without the need for nutrient supplementation, fully occupying the volume of vinasse with mycelium which thus became altogether a hard-solid mass. These results indicate that the ability of vinasse to kill M. perniciosa does not correspond to a generalized antifungal activity, and therefore depends on some specific trait of M. perniciosa biology.
Vinasse chemical composition is very complex [13]. Future approaches might allow the identification of the compounds or chemical groups in vinasse that are active against M. perniciosa, eventually enabling the creation of an engineered simpler and more efficient solution which considers the environmental effects in the long run. Moreover, the evaluation of the potential spectrum of vinasse application as a specific fungicide will have to be addressed experimentally. In particular, there is one possibility that stands out, which is that vinasse might affect the proliferation of M. roreri [29], another phytopathogenic filamentous fungus genetically very close to M. perniciosa also causing a severe and economically threatening disease in cacao fruits known as moniliasis or frosty pod rot [30].
![]()
M. perniciosa
Alta Mogiana
Batatais
Pedra
100%
50%
25%
100%
50%
25%
100%
50%
25%
CBS 441.80
-
-
+
-
+
+
-
+
+
CBS 442.80
-
-
-
-
-
+
-
+
+
Table 1: Growth (+) or absence of growth (-) of the M. perniciosa strains CBS 441.80 and CBS 442.80 when incubated in vinasse from the plants of Alta Mogiana, Batatais and Pedra, undiluted (100% vinasse) and in 2 different dilutions, 50% and 25% vinasse. Results presented were identical in three independent replicates.
Conclusions
Vinasse is the main waste product from sugarcane bioethanol production process. It has a very high fertilization ability due to its high N, P and K contents and has therefore been largely used to fertirrigate the sugarcane fields for decades. Nevertheless, if applied in excess it becomes a critical problem for soils and groundwater. Many suggestions in the literature regard solutions that promote the reduction of the amount of vinasse produced per litre of ethanol, its concentration, clearing or neutralization. Other suggestions regard the use of vinasse for biotechnological applications, including the growth of some economically interesting microorganisms or aerobic/anaerobic digestion. For now, the most cost-effective application for vinasse keeps being fertirrigation, in time improving soil quality and crop productivity [3,5], provided it is done carefully and responsibly, guaranteeing the control over all the environmental implications.
The utilization of vinasse has associated pollution problems that only recently are being recognized [3], but altogether it appears to be beneficial [6-9]. Our results clearly show the potential of vinasse to control the filamentous fungus M. perniciosa, cacao Witches’ Broom Disease causative agent. The impact of the introduction of vinasse in the cacao agro-ecosystem needs to be studied, not only in terms of chemical balances, but also in terms of the long-term effect on the associated soil microbiota. The experience from sugar-cane fields shows that the resident soil microbial community is very resilient [31,32], which comes into agreement with our finding that the inhibition of WBD by vinasse is not a generalized biocidal effect, which would be detrimental for the environment in the long run. The problem resides in the huge amounts of vinasse produced in Brazil every year, and the fact that this country was the most affected by the disease, the utilization of vinasse for its containment is an expressive possibility. There may be logistic and cost problems hindering the utilization of vinasse in the short-term deriving from the distances separating the bioethanol plants from the cacao fields, associated with this waste physicochemical properties, in particular its pH and corrosive nature. Nevertheless, these problems should in time be technically overcome in view of the advantage of contributing to solve two serious and urgent problems with a single procedure: introducing a new alternative for the disposal of high amounts of vinasse, reducing the serious danger to the environment and groundwater of unlawful discharges and precarious storage, and the containment of cacao Witches’ Broom Disease. Together, the procedure should impact positively in the overall economical equation and contribute significantly to the socio-economic recovery of the regions of Brazil that most suffered with the crisis caused by cacao production fall-out in the last decades.
Acknowledgements
We thank the researchers and technicians at Fermentec Lda, for kindly supplying the batches of vinasse from the three ethanol plants in Brazil, and sharing valuable information on vinasse and the industrial bio-ethanol production process. This work was supported by the strategic programme UID/BIA/04050/2013 (POCI-01-0145-FEDER-007569) funded by national funds through the FCT I.P. and by the ERDF through the COMPETE2020 - Programa Operacional Competitividade e Internacionalizacao (POCI), and the project EcoAgriFood (NORTE-01-0145-FEDER-000009) from Norte Portugal Regional Operational Programme (NORTE 2020) under the PORTUGAL 2020 Partnership Agreement through the European Regional Development Fund (ERDF). PF and MA-R are PhD students of the Doctoral Programmes in Applied and Environmental Microbiology (DP_AEM), (FCT grant PD/BD/113814/2015) and in Molecular and Environmental Biology (PDBMA), (FCT grant PD/BD/145354/2019), respectively.
References
- Carrilho ENVM, Labuto G, Kamogawa MY. Destination of Vinasse, a Residue from Alcohol Industry: Resource Recovery and Prevention of Pollution. In: Prasad MNV, Shih K, editors. Environmental Materials and Waste: Academic Press. 2016: 21-43.
- Lopes ML, Paulillo SC, Godoy A, Cherubin RA, Lorenzi MS, Giometti FH, et al. Ethanol production in Brazil: a bridge between science and industry. Brazilian Journal of Microbiology. 2016; 47: 64-76.
- Moran-Salazar RG, Sanchez-Lizarraga AL, Rodriguez-Campos J, Davila-Vazquez G, Marino-Marmolejo EN, Dendooven L, et al. Utilization of vinasses as soil amendment: consequences and perspectives. Springer Plus. 2016; 5: 1007.
- Naspolini BF, Machado ACO, Junior WBC, Guimaraaes FDN, Cammarota MC. Bioconversion of Sugarcane Vinasse into High-Added Value Products and Energy. BioMed Research International. 2017.
- Prado RM, Caione G, Campos CNS. Filter Cake and Vinasse as Fertilizers Contributing to Conservation Agriculture. Applied and Environmental Soil Science. 2013: Article ID 581984.
- Soobadar A, Ng Kee Kwong KFR. Impact of High Rates of Vinasse on Some Pertinent Soil Characteristics and on Sugarcane Yield in Mauritius. Journal of Sustainable Agriculture. 2012; 36: 36-53.
- Hazbavi Z, Sadeghi SHR. Potential effects of vinasse as a soil amendment to control runoff and soil loss. Soil. 2016; 2: 71-78.
- Bordonal RdO, Carvalho JLN, Lal R, de Figueiredo EB, de Oliveira BG, La Scala N. Sustainability of sugarcane production in Brazil. A review. Agronomy for Sustainable Development. 2018; 38: 13.
- Zani CF, Barneze AS, Robertson AD, Keith AM, Cerri CEP, McNamara NP, et al. Vinasse application and cessation of burning in sugarcane management can have positive impact on soil carbon stocks. Peer J. 2018; 6: e5398.
- Fuess LT, Rodrigues IJ, Garcia ML. Fertirrigation with sugarcane vinasse: Foreseeing potential impacts on soil and water resources through vinasse characterization. Journal of environmental science and health Part A, Toxic/hazardous substances & environmental engineering. 2017; 52: 1063-1072.
- Christofoletti CA, Escher JP, Correia JE, Marinho JFU, Fontanetti CS. Sugarcane vinasse: Environmental implications of its use. Waste Management. 2013; 33: 2752-2761.
- Botelho RG, Tornisielo VL, de Olinda RA, Maranho LA, Machado-Neto L. Acute toxicity of sugarcane vinasse to aquatic organisms before and after pH adjustment. Toxicological & Environmental Chemistry. 2012; 94: 2035-2045.
- Rodrigues Reis CE, Hu B. Vinasse from Sugarcane Ethanol Production: Better Treatment or Better Utilization? Frontiers in Energy Research. 2017; 5.
- Goldemberg J, Coelho ST, Guardabassi P. The sustainability of ethanol production from sugarcane. Energy Policy. 2008; 36: 2086-2097.
- Nitayavardhana S, Issarapayup K, Pavasant P, Khanal SK. Production of protein-rich fungal biomass in an airlift bioreactor using vinasse as substrate. Bioresource Technology. 2013; 133: 301-306.
- Money NP. Fungi and Biotechnology. In: Watkinson SC, Boddy L, Money NP, editors. The Fungi (Third Edition). Boston: Academic Press; 2016: 401-424.
- Gmoser R, Ferreira JA, Lundin M, Taherzadeh MJ, Lennartsson PR. Pigment Production by the Edible Filamentous Fungus Neurospora Intermedia. Fermentation. 2018; 4: 11.
- Nair RB, Taherzadeh MJ. Valorization of sugar-to-ethanol process waste vinasse: A novel biorefinery approach using edible ascomycetes filamentous fungi. Bioresource Technology. 2016; 221: 469-476.
- Aime MC, Phillips-Mora W. The causal agents of witches’ broom and frosty pod rot of cacao (chocolate, Theobroma cacao) form a new lineage of Marasmiaceae. Mycologia. 2005; 97: 1012-1022.
- Meinhardt LW, Rincones J, Bailey BA, Aime MC, Griffith GW, Zhang D, et al. Moniliophthora perniciosa, the causal agent of Witches Broom disease of cacao: what’s new from this old foe? Molecular Plant Pathology. 2008; 9: 577-588.
- Teixeira PJPL, Thomazella DPdT, Pereira GAG. Time for Chocolate: Current Understanding and New Perspectives on Cacao Witches’ Broom Disease Research. PLOS Pathogens. 2015; 11: e1005130.
- Marelli JP, Maximova SN, Gramacho KP, Kang S, Guiltinan MJ. Infection Biology of Moniliophthora perniciosa on Theobroma cacao and Alternate Solanaceous Hosts. Tropical Plant Biology. 2009; 2: 149-160.
- Pohlan HAJ, Pérez VD. Growth and Production of Cacao. In: Verheye WH, editor. Soils, Plant Growth and Crop Production - Volume III - Encyclopedia of Life Support Systems (EOLSS). Oxford, UK: EOLSS Publishers. 2010.
- Ferraz P, Cássio F, Lucas C. Potential of Yeasts as Biocontrol Agents of the Phytopathogen Causing Cacao Witches’ Broom Disease: Is Microbial Warfare a Solution? Frontiers in Microbiology. 2019; 10: 1766.
- Almeida OC, Chiacchio F, Rocha HM. Survival of Crinipellis perniciosa (Stahel) Singer on dried brooms of cacao (Theobroma cacao L.) in the State of Bahia (in Portuguese). Agrotrópica (Brazil). 1997; 9: 23-28.
- Medeiros FHV, Pomella AWV, de Souza JT, Niella GR, Valle R, Bateman RP, et al. A novel, integrated method for management of witches' broom disease in Cacao in Bahia, Brazil. Crop Protection. 2010; 29: 704-711.
- Weir BS, Johnston PR, Damm U. The Colletotrichum gloeosporioides species complex. Stud Mycol. 2012; 73: 115-180.
- Talhinhas P, Loureiro A, Oliveira H. Olive anthracnose: a yield and oil quality degrading disease caused by several species of Colletotrichum that differ in virulence, host preference and geographical distribution. Molecular Plant Pathology. 2018; 19: 1797-1807.
- Barbosa CS, Fonseca RRd, Batista TM, Barreto MA, Argolo CS, Carvalho MRd, et al. Genome sequence and effectorome of Moniliophthora perniciosa and Moniliophthora roreri subpopulations. BMC Genomics. 2018; 19: 509.
- Bailey BA, Evans HC, Phillips-Mora W, Ali SS, Meinhardt LW. Moniliophthora roreri, causal agent of cacao frosty pod rot. Molecular Plant Pathology. 2018; 19: 1580-1594.
- Lourenço KS, Suleiman AKA, Pijl A, Cantarella H, Kuramae EE. Dynamics and resilience of soil mycobiome under multiple organic and inorganic pulse disturbances. Science of the Total Environment. 2020; 733: 139173.
- Lourenço KS, Suleiman AKA, Pijl A, van Veen JA, Cantarella H, Kuramae EE. Resilience of the resident soil microbiome to organic and inorganic amendment disturbances and to temporary bacterial invasion. Microbiome. 2018; 6: 142.