
Research Article
Austin J Biotechnol Bioeng. 2015;2(3): 1046.
Bioengineering of Cow Metabolic Rhythms Through Feeding Timing
Akbar Nikkhah*
Department of Animal Sciences, University of Zanjan, Iran
*Corresponding author: Akbar Nikkhah, Chief Highly Distinguished Professor, Foremost Principal Highly Distinguished Elite-Generating Scientist, Department of Animal Sciences, Faculty of Agricultural Sciences, University of Zanjan, National Elite Foundation, Iran
Received: June 18, 2015; Accepted: July 07, 2015; Published: June 09, 2015
Abstract
The objective was to determine effects of feed presentation time and dietary forage to concentrate ratio on circadian and postprandial patterns of feed intake in lactating cows. Four multiparous and four primiparous Holstein cows were used in a 4 × 4 Latin square design with a 2 × 2 factorial arrangement of treatments. A higher or a lower concentrate Total Mixed Ration (TMR) was delivered at either 2100 h or 0900 h. The study consisted of four 21-d periods, each with 14-d of adaptation. A metabolic acquisition system was used to monitor continuous feed intake electronically. Feed presentation at 2100 h vs. 0900 h increased the amount of feed consumed within 3-h post-feeding from 26 to 37% of total daily intake. Dry Matter Intake (DMI) over the 24-h remained similar. Provision of TMR at 2100 h vs. 0900 h increased DMI in primiparous cows but not in multiparous cows. Results demonstrated that evening vs. morning feeding increased eating rate and feed intake shortly post-feeding. Timing of feeding, thus, could be a bioengineer and synchronizer of periprandial metabolic rhythms in lactating dairy cows.
Keywords: Intake rhythm; Feeding timing; Bioengineering; Dairy cow
Introduction
Fresh feed delivery determines circadian patterns of eating activity in lactating cows [1,2]. Effects of feed presentation timing on circadian and postprandial patterns of feed intake need to be quantified to enable accurate analysis of feeding behavior and prediction of feed intake. Robinson et al. [3] fed lactating cows 67% of the TMR at 0800 h and 33% of it at 1800 h, and fed a protein supplement (about 15% of estimated daily intake) at 0830 h or 0030 h. Presenting the protein supplement at 0030 h vs. 0830 h significantly increased protein meal Dry Matter Intake (DMI) and numerically increased total DMI (16.92 vs. 15.94 kg/d). The midnight compared to morning presentation of the protein supplement stabilized rumen fermentation. Restricting day-time feed access and shifting the eating time to evening hours reduced both DMI and energy expenditure [4]. Under cold weather, feed presentation at 2000 h instead of 0900 h improved growth rate in beef steers without affecting DMI [5]. Kennedy et al. [6] fed heifers at either 0900 or 2000 h in winter (<- 15°C) and found that evening instead of morning feed presentation improved feed efficiency in backgrounding period. In cows exposed to 5-d phases of thermoneutral (24°C during the day and 20°C overnight) and heat stress (32°C) conditions followed by a 5-d recovery period, evening feeding reduced heat-stress-phase DMI [7]. Altogether, DMI responses to feeding time have thus far been studied in terms of animal thermodynamics. It remains unknown if and how feed presentation time regulates feed intake and its postprandial patterns under thermoneutral conditions. The objective was to open a new horizon into ruminant chronophysiology by determining feeding time effects on intake periprandial and circadian patterns in lactating dairy cows.
Materials and Methods
Cow management and experimental design
Four multiparous (BW = 652 ± 14 kg, BCS = 2.87 ± 0.14, days in milk = 83 ± 22) and four primiparous (BW = 667 ± 110 kg, BCS = 3.19 ± 0.66, 81 ± 23 days in milk; mean ± SD) lactating Holstein cows were housed in tie stalls. All experimental procedures involving animals were according to the local guidelines of animal Care. The average air temperature and relative humidity were 20.4°C and 68.1%, respectively. The maximum air temperature of the metabolism unit did not exceed 25°C at any time during the experiment. Cows were offered either a higher concentrate diet (HC) with a forage to concentrate ratio (F:C) of 38.5:61.5 or a lower concentrate diet (LC) with a F:C of 50.6:49.4. The total mixed ration (TMR, Table 1) was delivered at either 0900 h or 2100 h. The experimental design was a replicated 4 × 4 Latin square with a 2 × 2 factorial arrangement of feed presentation time and dietary concentrate level. Each experimental period lasted for 21 d with 14 d of adaptation. Cows were fed ad libitum permitting 5-10% orts, with unlimited access to fresh water. Circadian and postprandial patterns of feed intake were monitored continuously using a data acquisition system within the metabolism unit. Total mixed rations were prepared every morning using a Data Ranger Mixer (American Calan, Northwood, NH) with a Weigh Tronix head (Model 1000, American Calan, Northwood, NH). Except for sampling weeks, cows were allowed for 2 h daily exercise (0700- 0900 h). Milking was performed twice daily in the stalls at 0400 and 1600 h. Lights were on from 0345 until 2245 h.
![]()
Experimental Diets
Diet ingredients
Higher Concentrate (HC)
Lower Concentrate (LC)
Alfalfa silage
15.87
20.96
Corn silage
22.67
29.66
Energy supplement
49.89
37.15
Protein supplement
11.57
12.23
Forage: concentrate ratio (F:C)
38.5 : 61.5
50.6 : 49.4
Crude protein %
18.1
17.3
Acid detergent insoluble protein %
1.5
1.6
Non-fiber carbohydrates %
39.7
35.9
Netural detergent fiber %
28.6
33.8
Acid detergent fiber %
15.1
19.4
Ether extract %
5.9
5.3
Table 1: Dietary forage and concentrate portions and chemical composition (DM basis).
Statistical analyses
The amount of feed consumed within eight 3-h intervals (24- h phase) were expressed as percent of total daily TMR intake for individual cows. The amount ingested within each 3-h interval was compared between evening-fed and morning-fed cows with Mixed Model Procedures of SAS program [8]. The effects of feeding time, diet, parity, and their interaction were fixed, and effects of period and cow within parity were random. Tukey’s multiple range test was used to compare the differences between least square means. Significant levels of fixed effects were declared at P ≤ 0.05 and trends were discussed at 0.05 < P ≤ 0.10.
Results and Discussion
Feed presentation time altered (P < 0.05) circadian and postprandial patterns of feed intake (Figure 1). The proportion of daily TMR intake consumed within 3-h post-feeding was 37% for 2100 h-fed cows and only 26% for 0900 h-fed cows (P < 0.05). In cumulative terms, the amounts consumed between 0-6 h and 0-9 h post-feeding were similar between the two groups (Figure 1). Parity and diet did not interact with feed presentation time on circadian patterns of feed intake (P > 0.10). Despite altering the post-feeding rhythms of intake, provision of TMR at 2100 h vs. 0900 h did not affect daily DMI.
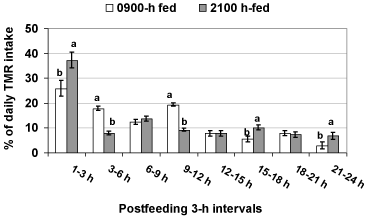
Figure 1: Post-feeding patterns of feed intake in cows fed at either 0900 h or
2100 h. Within each 3-h, different superscripts indicate P < 0.05.
Fresh feed delivery is a major determinant of circadian rhythms in feed intake of tie-stall-housed and loose-housed lactating cows [1,9]. The stimulatory effect of feed presentation on eating activity can persist even with four times daily feeding [10]. These authors reported that cows fed once daily at 0530 h spent less time eating than cows fed twice daily at 0530 and 1515 h. The longer eating time of cows fed twice daily was mostly due to increased eating time between 2000 to 0600 h. However, when Phillips and Rind [2] presented fresh TMR either once daily at 0600 h or four times daily at 0600, 1000, 1400, and 1900 h to early-lactation cows in free stalls, the total time spent eating did not differ. Nonetheless, cows fed four times daily tended to spend more time eating in evening (1600-2000 h) than in morning (0400-1200 h). The current data and the results of DeVries et al. [10] and Phillips and Rind [2] suggest that dairy cows eat when they are presented fresh feed and that the amount consumed after feed presentation depends on time of day. DeVries et al. [10] and Philips and Rind [2] did not quantify the proportional contribution of the more frequent feed delivery and 2) evening feed delivery per se to greater eating time. DeVries et al. [10] did not monitor daily DMI. Phillips and Rind [2] suggested that if cows anticipate the feed presentation, they would exhibit a more pronounced peak in time spent eating upon fresh feed delivery. Thus, the possibility exists that cows may have effectively anticipated the 2100 h-feed delivery in the current study. Plasma insulin was higher and plasma glucose was lower at 2 h post-feeding in 2100-fed cows than in 0900 h-fed cows [11]. Higher insulin could mean a weakened action of glucagon, thus reducing gluconeogenesis [12]. The intraperitoneal injection of glucagon antibodies to suppress glucagon effects stimulates feeding [13]. The intravenous glucagon has been shown to reduce feed intake in sheep [14,15]. Probably the higher blood insulin and lower blood glucose at 2 h post-feeding in evening-fed cows may have delayed the glucagon-driven satiety. This may have contributed to the increased eating rate within 3 h of the evening feed presentation. Moreover, anticipation of imminent ‘lights off’ might also induce eating activity.
Conclusion
Presenting higher and lower concentrate TMRs at 2100 h vs. 0900 h consistently increased eating rate and the feed amount consumed within 3-h post-feeding in primiparous and multiparous lactating dairy cows. The feed consumed within 3-h of feeding was increased from 26% to 37% of total daily intake. However, total daily DMI was similar. Results establish that evening compared to morning feed presentation increased feed intake and eating rate shortly post-feeding, and thereby altered circadian intake patterns in lactating cows. Certain times of feed presentation are synchronizers of postprandial intake patterns.
Acknowledgement
The Ministry of Science Research and Technology, National Elite Foundation, and University of Zanjan are thanked for supporting the author’s global initiatives and programs of optimizing science education in the third millennium.
References
- DeVries TJ, von Keyserlingk MA, Beauchemin KA. Short communication: Diurnal feeding pattern of lactating dairy cows. J Dairy Sci. 2003; 86: 4079-4082.
- Phillips CJ, Rind MI. The effects of frequency of feeding a total mixed ration on the production and behavior of dairy cows. J Dairy Sci. 2001; 84: 1979-1987.
- Robinson PH, Gill M, Kennelly JJ. Influence of time of feeding a protein meal on ruminal fermentation and forestomach digestion in dairy cows. Journal of Dairy Science. 1997; 80: 1366-1373.
- Aharoni Y, Brosh A, Harari Y. Night feeding for high-yielding dairy cows in hot weather: effects on intake, milk yield and energy expenditure. Livestock Production Science. 2005; 92: 207-219.
- Small JA, Kennedy AD, Veira DM, McCaughey WP, Ward DR. Time of feeding and growth promotant effects on the winter growth performance and carcass traits of steers. Canadian Journal of Animal Science. 2004; 84: 133-144.
- Kennedy AD, Bergen RD, Lawson TJ, Small JA, Veira DM. Effects of evening feeding and extended photoperiod on growth, feed efficiency, live animal carcass traits and plasma prolactin of beef heifers housed outdoors during two Manitoba winters. Canadian Journal of Animal Science. 2004; 84: 491-500.
- Ominski KH, Kennedy AD, Wittenberg KM, Moshtaghi Nia SA. Physiological and production responses to feeding schedule in lactating dairy cows exposed to short-term, moderate heat stress. J Dairy Sci. 2002; 85: 730-737.
- SAS Institute, Inc. SAS/STAT user’s guide. V. 9.1. SAS Institute, Inc., Cary, NC. 2003.
- Haley DB, Rushen J, de Passille AM. Behavioral indicators of cow comfort: Activity and resting behaviour of dairy cows housed in two types of housing. Canadian Journal of Animal Science. 2000; 80: 257–263.
- DeVries TJ, von Keyserlingk MA, Beauchemin KA. Frequency of feed delivery affects the behavior of lactating dairy cows. J Dairy Sci. 2005; 88: 3553-3562.
- Furedi C, Kennedy AD, Nikkhah A. Plaizier J. Glucose tolerance and diurnal variation of circulating insulin in evening and morning fed lactating cows. Advances in Dairy Technology. 2006; 18: 356.
- Brockman RP. Effect of insulin on the utilization of propionate in gluconeogenesis in sheep. Br J Nutr. 1990; 64: 95-101.
- Langhans W, Zeiger U, Scharrer E, Geary N. Stimulation of feeding in rats by intraperitoneal injection of antibodies to glucagon. Science. 1982; 218: 894-896.
- Deetz LE, Wangsness PJ. Influence of intrajugular administration of insulin, glucagon and propionate on voluntary feed intake of sheep. J Anim Sci. 1981; 53: 427-433.
- Canadian Council on Animal Care. Guide to the Care and Use of Experimental Animals. Olfert ED, Cross BM, McWilliam AA, editorss. CCAC, Ottawa, ON, Canada. 1993; 1.