
Special Article - Cataract Clinical Cases and Images
Austin J Clin Ophthalmol. 2016; 3(2): 1067.
Age Related or Senile Cataract: Pathology, Mechanism and Management
Sreelakshmi V and Abraham A*
Department of Biochemistry, University of Kerala, Kariavattom, India
*Corresponding author: Annie Abraham, Department of Biochemistry, University of Kerala, Kariavattom, Thiruvananthapuram, 695581, Kerala, India
Received: April 04, 2016; Accepted: June 09, 2016; Published: June 15, 2016
Abstract
Cataract is a serious eye disease accounts for the major cause of blindness globally. It is characterized by the loss of transparency and opacification of eye lens; an opaque lens scatters the light as it passes through it and prevents the sharpness of the image in the retina and vision becomes blurred. Cataractogenesis is associated with numerous factors acting over many years. The major reason lies behind the formation of cataract is the damage induced by free radicals, reactive oxygen/ nitrogen species to the crystalline lens. In this review, we have discussed the different events and mechanisms associated oxidative damage in the lens that gives rise to cataractogenesis, the present treatment procedures and management of cataract.
Keywords: Cataract; Eye; Lens; MAPK pathway; Oxidative stress
Introduction
The visual system is the various components of eyes functioning in the process of vision by reacting to light, gain information about their environments and help to recognize the outer world by the process of visual perception and the resulting perception is called vision or sight. Vision is one of the most complex functions and it requires the cooperation of many intricate parts and the eye is made up of three coats. The outer layer ortunica externa or tunica fibrosas composed of the cornea and sclera. The middle layer or tunica media or tunica vasculosa or uvea consists of the choroid, ciliary body and iris. The inner layer or tunica interna or tunica nervosa or retinais the light-sensitive tissue layer equipped with photoreceptors. Within these coats are the aqueous humour, the vitreous body and the flexible lens. The aqueous humour is a clear fluid that is contained in two areas: the anterior chamber between the cornea and the iris and the posterior chamber between the iris and the lens. The lens is suspended to the ciliary body by the suspensory ligament (Zonule of Zinn) made up of fine transparent fibers. The vitreous body is a clear jelly that is much larger than the aqueous humour present behind the lens, and the rest is bordered by the sclera, zonule and lens. Vision begins when light rays are reflected off an object and enter the eyes through the cornea, the transparent outer covering of the eye. The cornea bends or refracts the rays that pass through a round hole called the pupil. The iris, or colored portion of the eye that surrounds the pupil, opens and closes to regulate the amount of light passing through. The light rays then pass through the lens, which actually changes shape so it can further bend the rays and focus them on the retina at the back of the eye. The retina is a thin layer of tissue at the back of the eye that contains millions of tiny light-sensing nerve cells called rods and cones, for bright light and dim light respectively. These cells in the retina convert the light into electrical impulses. The optic nerve sends these impulses to the visual cortex in the brain where a composite image is produced [1].
The lens plays a crucial role in focusing unimpeded light on the retina. Eye lens is a biconvex, transparent, elastic, avascular structure that is located just behind the iris and the pupil that receives all its nutrients from aqueous and vitreous humor. The lens is suspended in place by the zonularfibres, which attach to the lens near its equatorial line and connect the lens to a ring of muscular tissue, called the ciliary body. Changing focus to an object at a greater distance requires the relaxation of the ciliary muscle, which in turn increases the tension on the zonules, flattening the lens and thus increasing the focal distance The lens is capable of changing its shape, functions to change the focal distance of the eye so that it can focus on objects at various distances, thus allowing a sharp real image of the object of interest to be formed on the retina by the process, accommodation [2]. The lens is flexible and its curvature is controlled by ciliary muscles through the zonules.
Structurally, the lens has three main components; capsule, epithelium and fibers. The capsule is the transparent, elastic, acellular basement membrane that completely encloses the whole cell mass and is the thickest basement membrane of the body. It is made up of type IV collagen and glycosaminoglycans and its main function is in the process oaccommodation bymolding the shape of the lens in response to tension from zonules [3]. The lens epithelium represents a single sheet of cuboidal cells just beneath the capsule at the anterior surface of the lens and the intercellular communication between the adjacent epithelial cells is through gap junctions. These monolayered cells regulate most of the homeostatic functions such as nutrient and ion transport, energy metabolism etc. in the lens and maintain the transparency of the lens. The fibers are long, thin, transparent cells form the bulk of the lens that the epithelial cells elongate, divide and differentiate to form the regularly arranged lens fibres. The new lens fibres are laid on the older deeper fibres and are formed throughout the life. Lens fibers arranged in zones, the cytoplasm of the cells of superficial bow region and the newly formed lens fibres contain nucleus, mitochondria, golgi complex, rough endoplasmic reticulum and polysomes and later on, all the light scattering organelles undergo an in built suicide process that minimizes light scatter and favors transparency [4].
Lens is an unusual organelle in its composition that with extraordinarily high protein content and low water content and this enables the lens to have a refractive index considerably greater than its fluid environment. Transparency of the lens is made possible by various factors such as normal physiology of epithelial cells, regular arrangement of the lens fibers, architecture of structural and functional proteins etc. Any alteration in the normal architecture of eye lens is associated with the change in the clarity of the lens or pacification and eventually forms the cataract. It is a significant visual impairment globally and as per the latest statistical records of World Health Organization (WHO), the total number of persons with visual impairment worldwide in 2010 was 285 million and cataract is responsible for 51% of world blindness, which represents about 20 million people [5]. Cataracts may be congenital, age related or secondary. Congenital cataracts, which are present at the birth and are, the less common cataract. The main types of age-related cataracts are nuclear sclerosis, cortical and posterior subcapsular. Nuclear cataracts form in the center of the lens and cause the nucleus to become hard or sclerotic with the deposition of brown pigment. Cortical cataracts are due to the opacity lens cortex and posterior subcapsular cataracts attack the back of the lens adjacent to the capsule. Secondary cataracts are caused by diseases like glaucoma and diabetes or medications such as steroids and radiations [6]. Cataract is associated with the gradual reduction of visual quality and is accompanied by a series of pathways that associated with imbalance in oxidant-antioxidant status [7], membrane lipid peroxidation [8], defected cellular communication [9], ion imbalance [10], modification, aggregation and accumulation of proteins [11,12], lenticular cell death [13,14] inflammation [15,16] etc. Hence, based on a variety of model systems; including cell/ organ culture, animal and human studies, the review focused on exploring the various pathways relating to the pathology of cataract, current treatment modalities and therapeutic preventive measures.
Mechanism of Cataract Formation
Oxidant-antioxidant imbalance
As lens is an organelle that exposed to light throughout the life time and prone to oxidative attack induced by reactive oxygen/ nitrogen species (ROS/RNS) [17,18], it is equipped with an efficient antioxidant system for defending these oxidative/nitrosative stress. The major enzymatic antioxidants in the lens are superoxide dismutase [19], Catalase [20], glutathione peroxidase [21], glutathione reductase, glutathione-S-transferase [22], thioredoxin system etc. [23]and non-enzymatic antioxidants are reduced glutathione [24], ascorbic acid, Vitamin A, E etc. [25-28]. These antioxidants protect lens from damage induced by toxic radicals/species and oxidative stress is a metabolic state in which excessive levels of highly reactive and unstable compounds overwhelm the ability of antioxidants that quenches them. Decline in the activity of all these enzymes and molecules are reported in the formation of cataract [29,30].
Stress signaling
NFκB is a ubiquitous transcription factor activated by ROS. Normally it is located in the cytoplasm in an inactive complex with inhibitor kappa B (Iκ B) and oxidative stress induce the release of I κ B resulting in translocation of NFκB to the nucleus and it binds to DNA control elements and thus influences the transcription of specific genes associated with stress signaling and cell death. NFκBmediated pathway is reported to present in lens epithelial cells exposed to hydrogen peroxide [31] and UV stress [32] indicating its role incataractogenesis.
MAPK pathway
Mitogen-activated protein kinases (MAPKs) are serine-threonine protein kinases that play the major role in the regulation of cell proliferation, cell differentiation and cell death. MAPKs family is characterized by the conserved activation domain and specialized activation module and it comprised of extracellular signal-regulated kinases (ERK-1 and ERK-2 isoforms), the c-Jun N-terminal kinases (JNK-1, JNK-2, and JNK-3 isoforms) and the p38 MAPKs (p38α, p38β, p38γ and p38d isoforms). Each subgroup of MAPKs is activated through a cascade of sequential phosphorylation events, beginning with the activation of MAPK kinase kinases (MAP3Ks). The MAP3Ks in turn phosphorylate and activate downstream MAPK kinases (MAP2Ks), which in turn stimulate MAPK activity through dual phosphorylation on threonine and tyrosine residues within a conserved tri-peptide motif. Activated MAPKs phosphorylate diverse substrates in the cytosol and nucleus to bring about changes in protein function and gene expression that execute the appropriate biological such as proliferation, differentiation, inflammatory responses, apoptosis etc. (Figure 1). MAPK phosphatases (MKPs), which recognize the TXY amino acid motif present in MAPKs, dephosphorylate and deactivate MAPKs [33]. MAPK pathways play discrete roles in the survival and normal functioning of lenticular epithelial cells and thus the transparency of the lens [34]. Oxidative stress is a predominant extracellular stimulus that activates MAPK pathways and many reports confirms the involvement of MAPK pathway in lens epithelial cell death and cataract formation through the disorganization of gap junctions and cytoskeletal assembly in the lens [35-37].
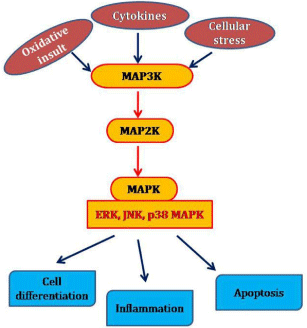
Figure 1: Schematic representation of MAPK Pathway.
Protein kinase pathway
Protein kinase C (PKC) is a family of serine/threonine kinases that functions in the process of cellular signaling by phosphorylation and like MAPKs, PKCs are activated by oxidant damage and calcium. The PKCs have been grouped into three classes (PKC α,β and γ) depending on the cofactors required for their activation. PKCγ moves to the plasma membrane following activation and phosphorylates targets such as receptors, structural proteins and gap junction proteins and reported in lens opacity [38-40].
Protein modification and removal of modified proteins
Proper arrangement of lens proteins plays the major role in the maintenance of transparency and modification of structural and functional proteins in the lens as a result of oxidation, proteolysis, transamidation, carbamylation, phosphorylation etc. is reported in the lens opacity [41].
Xanthine oxidase is a pro-oxidant enzyme normally present in ocular tissues and the raised activity of xanthine oxidase forms an important source of free radicals and is a lens oxidative stress marker [42]. Oxidants generated by xanthine oxidase system imposes crosslinking and aggregation of crystallins, loss of cellular redox balance, oxidation of DNA bases and lipid peroxidation of polyunsaturated fatty acids and inflict the damage to the lens membrane active transport pumps and implicated in cataractogenesis [43]. Proteins may contain several actual or potential sulfhydryl groups and are the potential sites for reversible oxidation-reduction reactions and thioltransferases in the lens prevent the oxidation of proteins and maintain redox homeostasis in the lens. Modification of proteins precedes a cascade of events starting with protein disulfide cross-links, alteration of protein conformation, protein/enzyme deactivation, protein solubility loss and eventually lens opacification [11,44].
Carbonylation of proteins is a metal accelerated modification of proteins occurring in the side chain of amino acids such as lysine, arginine, proline or histidine and is a widespread indicator of severe oxidative damage an irreversible oxidative damage leading to the loss of protein function. carbonylated proteins tend to form high molecular-weight aggregates that are resistant to degradation, accumulate as damaged or unfolded proteins and carbonyls are reported in the lenses of cataractous lenses [45].
Crystallins are the predominant structural proteins in the lens constitute about 90% of water soluble proteins of the lens and add to the transparency and refractive properties by its proper packaging. The three main crystallin types found in the lens are α, β, and γ crystallins. Crystallins tend to form soluble, high molecular weight aggregates that pack tightly in lens fibres, thus increasing the index of refraction of the lens while maintaining its transparency. α crystallinsare the major crystallins comprised of α A and α B crystallinshave chaperone function [46,47] involved in remodeling and protection of the cytoskeleton, inhibition of apoptosis and resistance towards oxidative stress [48] (Andley 2007). Also, the molecular association of β and γ crystallins form dense packaging and this minimizing light scattering and provides an optimum in the transparency of lens [49]. Several studies have described extensive truncation of human lens α, β, and γ Crystallins duringcataract development through several modifications such as aggregation and cross-linking which leads toprotein insolubilization [50,51].
Intermediate filaments are the major cytoskeletal elements responsible for the transparency of the lens and the main intermediate filaments are filensin, phakinin and phakinin. Filensin and phakinin forms complex with αA and αB crystallins and they together termed as beaded filaments and vimentinforms a network in the lens that functions in the lens architecture and also interacts with flensing [52]. Degradation of filensin and phakinin and vimentindeamidation was reported in the age-related cataract lenses [53,54].
Gap junctions are transmembrane aqueous channels that connect the cytoplasm of neighboring cells and allow the passage of molecules up to the size of 1 kDa between the connected cells and they respond to a variety of factors, such as Ca2+ levels, voltage, pH and phosphorylation events [55,56]. Different arrangements of gap junctions exist between the different regions of epithelial cells, between the fiber cells, and between the epithelial and fiber cells [57]. Connexins are a family of four-pass transmembrane proteins that assemble in groups of six to form hemichannels or connexons and two hemichannels then combine to form a gap junction. The combination of different connexins differs in the physiological properties, including unitary conductance, permeability, gating, and regulation by different protein kinase-dependent pathways. Connexins play the major role in the lens transparency and cataract that they maintain the intracellular symphony in the lens. With defective hemichannels and gap junction, metabolite supply and catabolite removal reduced the metabolic center of the lens; epithelial cells become malfunctioned resulting in vision impairment. Connexins 43, 46 and 50 are the major connexins in the lens although their distribution varies among lens epithelia and fibers. Although connexins are phosphoproteins, many abnormal phosphorylation patterns of connexins are reported in opaque lenses. All the connexin gap junctions and hemichannel functions are affected by oxidative stress and implicated in the formation of cataract [58]. Connexins are prone to phosphorylation by PKCs and nitrosylation by nitric oxide [59]. PKC-dependent phosphorylation of Ser368 in Cx43 affects its channel behavior, which leads to the decreased intercellular communication and altered permeability [60,61].
Transglutaminase 2 is an inducible transamidatingacyltransferase that catalyzes Ca2+ dependent protein modifications by inducing of covalent cross-links between peptide bound glutamine and lysine residues and is up-regulated often in cells undergoing oxidative stress and apoptosis [62,63]. Deamidation and further denaturation of crystallinsare implicated in the pathology of cataract [64,65]. Deamidation causes unfolding of α Crystallin molecules; partially unfolded protein may possess increased exposure to key hydrophobic residues, which are involved in the chaperone activity [66].
Matrix metalloproteinases (MMPs) represent a family of endopeptidases that are capable of degrading the extracellular matrix molecules and thereby of maintaining normal physiological processes such as morphogenesis and influencing cell biological activities [67] and the most widely studied MMPs in the ocular tissues are MMP 2 and MMP 9. Lens cells are capable of synthesizing MMPs upon injury to the lens, such as UV irradiation [68], oxidative stress [69] and MMPs digests type IV collagen and laminin, cleaves and aggregates βB1 crystallinand produce lens opacity [15,70].
Oxidant damage to lens proteins and its accumulation is implicated in the formation of cataract. For maintaining the transparency of lens, the damaged proteins should be removed and the process is done by the ubiquitin-proteasome mechanism. Degradation of a protein via the ubiquitin-proteasome pathway involves tagging of the substrate protein by the covalent attachment of multiple ubiquitin molecules and the subsequent degradation of the tagged protein by the 26S proteasome. Multiple enzymes are involved in the process of ubiquitinylation through a series of ATP-dependent enzymatic steps. In this process, ubiquitin is first activated by ubiquitin activating enzyme (E1), activated ubiquitin is then passed to a ubiquitin carrier protein (E2) both via the formation of a thiol ester bond. The activated ubiquitin is then either directly linked to substrates or is linked to substrates via ubiquitin ligase (E3), targeted to 26S proteasome for proteolysis and ubiquitin is recycled [71]. Ubiquitin-dependent proteasome pathway is under the control of oxidant response and the activity of both activating and conjugating enzymes were altered in the aged lens [72,73] and amino acid residues of ubiquitin which is involved in the protein degradation mechanism is modified by oxidants [74]. All these events compromise the function of ubiquitinproteasome pathway in the cataractous lens.
Lipid peroxidation
The integrity of lenticular membrane is one of the important factors that keep lens transparency. Lens membrane serves as an impermeable barrier to cations and is equipped with membrane ATPases for the homeostasis of water, calcium, sodium and potassium. Lipid oxidation is a major harmful consequence of ROS formation as it produces irreversible oxidative changes of membranes. It has been shown to induce disorganization of the membrane, modification of membrane proteins, alters the physiological functions of cell membranes. Lipid peroxidation has been implicated in the pathogenesis of cataract because the lethal peroxidation products induce fragmentation of soluble lens proteins and damage critical membrane structures, epithelial cell apoptosis and correlating with an increase in lens opacity and changes in the refractive properties of the lens [8,75-77].
Ion imbalance
As discussed earlier, lens membrane is equipped with various pumps for maintaining the optimal ion homeostasis. To maintain such steep ionic gradients, the lens must continually expend energy to drive sodium and calcium outward, at the same time functioning to accumulate other ions such as potassium. Cellular calcium homeostasis is achieved by a balance between the inward leak and out flow by plasma membrane Ca2+ ATPase and Na+ Ca2+ exchanger. Progressive elevation of sodium, marked loss of potassium and several fold increment of calcium are documented in the literature in the pathology of lens opacification [78,79]. A prolonged increase in the calcium concentration would be expected to activate proteases such as calpain and could induce the formation of protein aggregates and irreversible breakdown of important structural proteins andlead to lens opacification [80-82]. The decrease of calcium ATPase activity is also reported with cataract due to lipid structural changes [83,84], increase in the oxidation of sulf-hydrul groups [85] and downregulated ATP utilization [86].
The redox state of the cell is largely reliant on the trace elements iron and copper by virtue of their capability to go through reversible redox reactions and is maintained within strict physiological limits. Accumulation of these metals may disrupt the intracellular redox status, alter protein conformation and inhibit protein function through metal substitution and interactions with sulfhydryl groups by catalyzing the formation of reactive hydroxyl radicals via Fenton reaction [87]. Iron and copper is implicated in the pathogenesis of cataracts by damaging lipid membranes and lens capsule, crosslinking and insolubilization of lens proteins, leakage of beta and gamma crystallins into the aqueous humor through the production of hydroxyl radicals and peroxyl radicals [88-91].
Inflammation
Inflammation is a series complex biological response of body tissues to harmful stimuli; although Inflammation is a protective response, it is reported in various diseases. Nitric oxide (NO) is a free radical gas, signaling messenger and the role of the NO in inflammation are well established. Nitric oxide is normally present at a low concentration in the aqueous humor that bathes the lens [92]. Constitutive levels of NO production contribute normal ocular function, but in response to induction of inducible nitric oxide synthase (iNOS) by oxidant trauma, the production of NO is elevated. NO contribute to oxidation stress by developing more powerful oxidative agents suchas peroxynitrite with superoxide [93] (Horton, 2003) which is highly reactive and inflicts cytotoxicity and nitrosative stress to proteins [94] and reported in cataractogenesisby type IV collagen and laminin and cleaves βB1 crystallin [18,95].
Interleukin (IL) -18 is a pleiotropic cytokine belonging to the IL-1 family induces interferon (IFN) γ, NFκB and iNOS and plays an important role in inflammatory action [96,97]. The increased generation of ROS accelerates the production of inflammatory cytokines like IFN γ in the lens through MAPK pathway. It has been reported that IFN-g leads to cataract development by causing the apoptosis of lens epithelial cells and associated with cataract development [14,98].
Lens epithelial cell apoptosis
As discussed earlier, metabolic homeostasis of a single layer of lens epithelial cells is the critical event in maintaining the transparency of the entire lens [99]. The cells have a relatively long life span under normal physiological conditions, the factors such as oxidative stress alter the viability of lenticular epithelia resulting in lens opacification. Enormous number researches confirmed the role of lens epithelial cell death as the key biochemical event underlying the process of cataractogenesis through a series of events mentioned above [14,100,101]. Oxidative damage, increased calcium level, membrane damage, inflammatory responses etc. activate lenticular apoptosis through the activation of pro-apoptotic factors, Caspases and inhibition of anti-apoptotic agents [102].
Treatment of cataract
In the modern century, surgical removal of cloudy/opaque lens and replacement with a synthetic intraocular lens is the only available treatment for cataract. Phacoemulsification, extracapsular cataract extraction and intracapsular cataract extraction are the major surgical procedures employed for cataract treatment throughout the world [103].
Phacoemulsification
It is the most common procedure used by developed countries. In this technique, a very small incision (2-3 mm) is produced on the surface of the eye in or near the cornea. A thin ultrasound probe is inserted into the eye and uses ultrasonic vibrations (40,000 Hz) to dissolve (phacoemulsify) the clouded lens. These tiny fragmented pieces are then suctioned out through the same ultrasound probe. Once the cataract is removed, an artificial lens is placed into the thin capsular bag that the cataract previously occupied. In phacoemulsification, cataract surgery can perform in less than 30 minutes and usually requires only minimal sedation and no stitches are used to close the wound [104].
Extracapsular cataract extraction
This technique requires a larger incision so that the cataract can be removed in one piece without being fragmented inside the eye. It involves the removal of almost the entire natural lens through a large (usually 10-12 mm) incision made in the cornea or sclera, while the elastic lens capsule (posterior capsule) is left intact to allow implantation of an intraocular lens. This surgical technique requires a various number of sutures to close the larger wound and visual recovery is often slower.Manual small incision cataract surgery is an evolution extracapsular cataract extraction characterized by an appropriately constructed scleral tunnel that does not require suturing. Here the wound being relatively smaller than the above still markedly larger than a phaco wound [105].
Intracapsular cataract extraction
It involves the removal of the lens and the surrounding lens capsule in one piece and the intraocular lens is placed in front of the iris. The procedure has a relatively high rate of complications due to the large incision required and pressure placed on the vitreous body.
Complications of cataract surgery
Although surgery is an effective measure for cataract blindness, it is not free from post-operative complications. The major risks associated with post-surgery are inflammation (swelling and redness) in the eye, swelling of the retina (cystoid macular oedema); where fluid builds up between layers of the retina at the back of the eye, swelling of the cornea- where fluid builds up in the cornea at the front of the eye; this usually clears itself, retinal detachment; a rare complication where the retina (layer of nerve cells inside the back of the eye) becomes separated from the inner wall of the eye, infection in the eye, such as endophthalmitis (a rare bacterial infection), glaucoma, secondary cataract etc. [106].
Preventive protection of cataract
Even though cataract surgical techniques are efficient treatment modalities, there are people around the globe with barriers for accessing all these facilities because of insufficient financial resources, lack of awareness etc. [107]. As cataract is a protein degenerative disorder, its irreversible nature directs vision researchers in the track of preventive protection for the management of cataract blindness. A noteworthy vision researches indicate the role of nutritional intrusion as a way to reduce the risk of cataract. As oxidative stress forms the key behind the pathological events associated with cataract, many studies have been focusing to explore the efficacy of antioxidants in the prevention of cataract. Studies recommend that optimizing intakes of lutein, zeaxanthin, B vitamins, and multivitamin supplements may be advantageous in diminishing risk of nuclear and possibly cortical cataract [108]. Also many indigenous plants and plant-derived compounds have been shown to protect lens transparency in both in vitro and in vivo models such as lupeol, luteolin by modulating antioxidant status and preventing apoptosis [83], rutin through blocking chaperon activity of α Crystallin [109,110], curcuminby inhibiting iNOS [111], resveratolby lowering lipid peroxidation [112] etc. The plants reported to have anticataractogenic potential are Cassia tora [63,77], Vitexnegundo [113], Moringaoleifera [114], Ginkgo biloba [115], Ocimum sanctum [116] etc. Thus the incorporation of these nutrients, plants and compounds is recommended for eye health and to prevent the onset and maturation of lenticular opacities through the way of prevention by functional foods.
Conclusion
Thus, the pathophysiological position of the oxidant-antioxidant balancing systems in the lens and cataract blindness is justified here. Through MAPK pathway, oxidative damage on lens induces membrane damage, metal accumulation, protein modification and accumulation, inflammation, lenticular apoptosis, etc. (Figure 2) and all these alter the refractive properties of the lens resulting in the opacity and cataract. Although modern surgical procedures are available for the cataract treatment, it has its own limitations and complications and natural product based nutritional therapy is a newly emerging cost effective area in the field of vision research for cataract management by the way of precautionary protection.
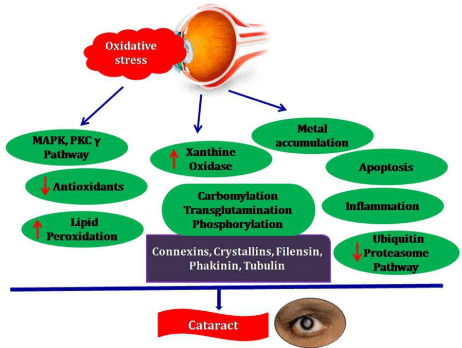
Figure 2: Schematic representation of events associated with
cataractogenesis.
Acknowledgement
Acknowledged to University Grants Commission (UGC), New Delhi for the financial support as a research grant.
References
- Sembulingam K, Sembulingam P. Essentials of Medical Physiology. New Delhi: Jaypeebrothers medical publishers Pvt Ltd. 2012.
- Forrester J, Dick A, McMenamin P, Lee W. The Eye: Basic Sciences in Practice. London: W. B. Saunders Company Ltd. 1996.
- Streeten BW. The nature of the ocular zonule. Trans Am Ophthalmol Soc. 1982; 80: 823-854.
- Hogan MJ, Alvarado JA, Weddell JE. Histology of the human eye. Philadelphia: WB Saunders. 1971.
- Pascolini D, Mariotti SP. Global estimates of visual impairment: 2010. Br J Ophthalmol. 2012; 96: 614-618.
- Yanoff M, Duker JS Ophthalmology. 3rd edn. Mosby Inc: St. Louis. 2008.
- Adedapo KS, Kareem ST, Bekibele CO, Nwobi NL. Increased oxidative stress and non enzymatic antioxidant levels in senile cataract. Arch ApplSci Res. 2012; 4: 2461-2466.
- Kisic B, Miric D, Zoric L, Dragojevic I, Stolic A. Role of lipid peroxidation in pathogenesis of senile cataract. Vojnosanit Pregl. 2009; 66: 371-375.
- Gao J, Sun X, Martinez-Wittinghan FJ, Gong X, White TW, Mathias RT. Connections between connexins, calcium, and cataracts in the lens. J Gen Physiol. 2004; 124: 289-300.
- Duncan G, Bushell AR. Ion analyses of human cataractous lenses. Exp Eye Res. 1975; 20: 223-230.
- Takemoto LJ. Oxidation of cysteine residues from alpha-A crystallin during cataractogenesis of the human lens. Biochem Biophys Res Commun. 1996; 223: 216-220.
- Boros S, Wilmarth PA, Kamps B, De Jong WW, Bloemendal H, Lampi K, et al. Tissue transglutaminase catalyzes the deamidation of glutamines in lens ßB2 and ßB3 crystallins. Exp Eye Res. 2008; 86: 383-393.
- Harocopos GJ, Alvares KM, Kolker AE, Beebe DC. Human age-related cataract and lens epithelial cell death. Invest Ophthalmol Vis Sci. 1998; 39: 2696-2706.
- Li WC, Kuszak JR, Dunn K, Wang RR, Ma W, Wang GM, et al. Lens epithelial cell apoptosis appears to be a common cellular basis for non congenital cataract development in humans and animals. J Cell Biol. 1995; 130: 169-181.
- Descamps FJ, Martens E, Proost P, Starckx S, Van den Steen PE, Damme JV, et al. Gelatinase B/matrix metalloproteinase-9 provokes cataract by cleaving lens ßB1 crystallin. FASEB J. 2005; 19: 29-35.
- Klein BE, Klein R, Lee KE, Knudtson MD, Tsai MY. Markers of inflammation, vascular endothelial dysfunction, and age-related cataract. Am J Ophthalmol. 2006; 141: 116-122.
- Babizhayev MA, Deyev AI, Linberg LF. Lipid peroxidation as a possible cause of cataract. Mech Ageing Dev. 1988; 44: 69-89.
- Inomata M, Hayashi M, Shumiya S, Kawashima S, Ito Y. Involvement of inducible nitric oxide synthase in cataract formation in Shumiya cataract rat (SCR). Curr Eye Res. 2001; 23: 307-311.
- Varma SD, Ets TK, Richards RD. Protection against superoxide radicals in rat lens. Ophthalmol Res. 1977; 9: 875-880.
- Bhuyan KC, Bhuyan DK. Catalase in ocular tissue and its intracellular distribution in corneal epithelium. Am J Ophthalmol. 1970; 69: 147-153.
- Pirie A. Glutathione peroxidase in lens and a source of hydrogen peroxide in aqueous humour. Biochem J. 1965; 96: 244-253.
- Rao GN, Sadasivudu B, Cotlier E. Studies on glutathione S-transferase, glutathione peroxidase and glutathione reductase in human normal and cataractous lenses. Ophthalmic Res. 1983;15: 173-179.
- Yegorova S, Yegorov O, Lou MF. Thioredoxin induced antioxidant gene expressions in human lens epithelial cells. Exp Eye Res. 2006; 83: 783-792.
- Giblin FJ. Glutathione: a vital lens antioxidant. J Ocul Pharmacol Ther. 2000; 16: 121-135.
- Heath H. The distribution and possible functions of ascorbic acid in the eye. Exp Eye Res. 1962; 1: 362-367.
- Jacques PF, Hartz SC, Chylack LT Jr, McGandy RB, Sadowski JA. Nutritional status in persons with and without senile cataract: blood vitamin and mineral levels. Am J Clin Nutr. 1988; 48: 152-158.
- Yeum KJ, Taylor A, Tang G, Russell RM. Measurement of carotenoids, retinoids, and tocopherols in human lenses. Invest Ophthalmol Vis Sci. 1995; 36: 2756-2761.
- Bernstein PS, Khachik F, Carvalho LS, Muir GJ, Zhao DY, Katz NB. Identification and quantitation of carotenoids and their metabolites in the tissues of the human eye. Exp Eye Res. 2001; 72: 215-223.
- Kisic B, Miric D, Zoric L, Ilic A, Dragojevic I. Antioxidant capacity of lenses with age-related cataract. Oxid Med Cell Longev. 2012; 2012: 467130.
- Stamenkovic M, Radic T, Stefanovic I, Coric V, Sencanic I, Pljesa-Ercegovac M, et al. Glutathione S transferase omega-2 polymorphism-Asn142Asp modifies the risk of age related cataract in smokers and subjects exposed to ultraviolet irradiation. Clin Exp Ophthalmol. 2014; 42: 277-283.
- Dudek EJ, Shang F, Taylor A. H(2)O(2)-mediated oxidative stress activates NF-kappa B in lens epithelial cells. Free Radic Biol Med. 2001; 31: 651-658.
- Boileau TW, Bray TM, Bomser JA. Ultraviolet radiation modulates nuclear factor kappa B activation in human lens epithelial cells. J Biochem Mol Toxicol. 2003; 17: 108-113.
- Morrison DK. MAP Kinase Pathways. Cold Spring HarbPerspect Biol. 2012; 4: 1-5.
- Li DW, Liu JP, Wang J, Mao YW, Hou LH. Expression and activity of the signaling molecules for mitogen-activated protein kinase pathways in human, bovine, and rat lenses. Invest Ophthalmol Vis Sci. 2003; 44: 5277-5286.
- Gong X, Wang X, Han J, Niesman I, Huang Q, Horwitz J. Development of cataractous macrophthalmia in mice expressing an active MEK1 in the lens. Invest Ophthalmol Vis Sci. 2001; 42: 539-548.
- Zhou J, Menko AS. Coordinate signaling by Src and p38 kinases in the induction of cortical cataracts. Invest Ophthalmol Vis Sci. 2004; 45: 2314-2323.
- Zhou J, Leonard M, Van Bockstaele E, Menko AS. Mechanism of Src kinase induction of cortical cataract following exposure to stress: destabilization of cell-cell junctions. Mol Vis. 2007; 13: 1298-1310.
- Lin D, Lobell S, Jewell A, Takemoto DJ. Differential phosphorylation of connexin46 and connexin50 by H2O2 activation of protein kinase Cgamma. Mol Vis. 2004; 10: 688-695.
- Zampighi GA, Planells AM, Lin D, Takemoto D. Regulation of lens cell-to-cell communication by activation of PKCgamma and disassembly of Cx50 channels. Invest Ophthalmol Vis Sci. 2005; 46: 3247-3255.
- Lin D, Takemoto DJ. Oxidative activation of protein kinase Cgamma through the C1 domain. Effects on gap junctions. J Biol Chem. 2005; 280: 13682-13693.
- Sharma KK, Santhoshkumar P. Lens aging: effects of crystallins. Biochim Biophys Acta. 2009; 1790: 1095-1108.
- Miric DJ, Kisic BB, Zoric LD. Mitic RV, Miric BM, Dragojevic IM. Xanthine oxidase and lens oxidative stress markers in diabetic and senile cataract patients. J Diabetes Complications. 2013; 27: 171-176.
- Varma SD, Hegde K, Henein M. Oxidative damage to mouse lens in culture. Protective effect of pyruvate. Biochim Biophys Acta. 2003; 1621: 246-252.
- Lou MF. Thiol regulation in the lens. J Ocul Pharmacol Ther. 2000; 16: 137-148.
- Balog Z, Klepac R, Sikic J, Jukic-Lesina T. Protein carbonylation and glycation in human lenses. Coll Antropol. 2001; 25: 145-148.
- Horwitz J. Alpha-crystallin. Exp Eye Res. 2003; 76: 145-153.
- Sun Y, MacRae TH. Small heat shock proteins: molecular structure and chaperone function. Cell Mol Life Sci. 2005; 62: 2460-2476.
- Andley UP. Crystallins in the eye: Function and pathology. Prog Retin Eye Res. 2007; 26: 78-98.
- Graw J. Mouse models of cataract. J Genet. 2009; 88: 469-486.
- Lampi KJ, Ma Z, Hanson SR, Azuma M, Shih M, Shearer TR, et al. Age-related changes in human lens crystallins identified by two-dimensional electrophoresis and mass spectrometry. Exp Eye Res. 1998; 67: 31-43.
- Srivastava OP, Srivastava K, Harrington V. Age-related degradation of betaA3/A1-crystallin in human lenses. Biochem Biophys Res Commun. 1999; 258: 632-638.
- Gounaria F, Karagiannia N, Minchevab A, Lichterb P, Georgatosc SD, Schirrmacher V. The mouse filensin gene: structure and evolutionary relation to other intermediate filament genes. FEBS Lett. 1997; 413: 371-378.
- Sanderson J, Marcantonio JM, Duncan G. A human lens model of cortical cataract: Ca2+-induced protein loss, vimentin cleavage and opacification. Invest Ophthalmol Vis Sci. 2000; 41: 2255-2261.
- Oka M, Kudo H, Sugama N, Asami Y, Takehana M. The function of filensin and phakinin in lens transparency. Mol Vis. 2008; 14: 815-822.
- Goldberg GS, Valiunas V, Brink PR. Selective permeability of gap junction channels. Biochim Biophys Acta. 2004; 1662: 96-101.
- Weber PA, Chang HC, Spaeth KE, Nitsche JM, Nicholson BJ. The permeability of gap junction channels to probes of different size is dependent on connexin composition andpermeant-pore affinities. Biophys J. 2004; 87: 958-973.
- Berthoud VM, Beyer EC. Oxidative stress, lens gap junctions, and cataracts. Antioxid Redox Signal. 2009; 11: 339-353.
- Retamal MA, León-Paravic CG, Verdugo CA, Alcaino CA, Amaro RM, Stehberg J. Connexin in lens physiology and cataract formation. J Clin Exp Ophthalmol. 2011; 1: 1-13.
- Bao L, Sachs F, Dahl G. Connexins are mechanosensitive. Am J Physiol Cell Physiol. 2004; 287: C1389-1395.
- Lampe PD, TenBroek EM, Burt JM, Kurata WE, Johnson RG, Lau AF. Phosphorylation of connexin43 on serine368 by protein kinase C regulates gap junctional communication. J Cell Biol. 2000; 149: 1503-1512.
- Ek-Vitorin JF, King TJ, Heyman NS, Lampe PD, Burt JM. Selectivity of connexin 43 channels is regulated through protein kinase C-dependent phosphorylation. Circ Res. 2006; 98: 1498-1505.
- Park D, Choi SS, Ha KS. Transglutaminase 2: a multi-functional protein in multiple subcellular compartments. Amino Acids. 2010; 39: 619-631.
- Sreelakshmi V, Abraham A. Anthraquinones and flavonoids of Cassia tora leaves ameliorate sodium selenite induced cataractogenesis in neonatal rats. Food Funct. 2016; 7: 1087-1095.
- Lorand L, Hsu LK, Siefring GE Jr, Rafferty NS. Lens transglutaminase and cataract formation. Proc Natl Acad Sci U S A. 1981; 78: 1356-1360.
- Hidasi V, Muszbek L. Transglutaminase activity in normal human lenses and in senile cataracts. Ann Clin Lab Sci. 1995; 25: 236-240.
- Smith JB, Liu Y, Smith DL. Identification of possible regions of chaperone activity in lens alpha-crystallin. Exp Eye Res. 1996; 63: 125-128.
- Sivak JM, Fini ME. MMPs in the eye: emerging roles for matrix metalloproteinases in ocular physiology. Prog Retin Eye Res. 2002; 21: 1-14.
- Sachdev NH, Di Girolamo N, Nolan TM, McCluskey PJ, Wakefield D, Coroneo MT. Matrix metalloproteinases and tissue inhibitors of matrix metalloproteinases in the human lens: Implications for cortical cataract formation. Invest Ophthalmol Vis Sci. 2004; 45: 4075-4082.
- Tamiya S, Wormstone IM, Marcantonio JM, Gavrilovic J, Duncan G. Induction of matrix metalloproteinases 2 and 9 following stress to the lens. Exp Eye Res. 2000; 71: 591-597.
- Alapure BV, Praveen MR, Gajjar D, Vasavada AR, Rajkumar S, Johar K. Matrix metalloproteinase-9 activity in human lens epithelial cells of cortical, posterior subcapsular and nuclear cataracts. J Cataract Refract Surg. 2008; 34, 2063-2067.
- Hershko A, Ciechanover A, Varshavsky A. Basic Medical Research Award. The ubiquitin system. Nat Med. 2000; 6: 1073-1081.
- Shang F, Gong X, Palmer HJ, Nowell TR Jr, Taylor A. Age-related decline in ubiquitin conjugation in response to oxidative stress in the lens. Exp Eye Res. 1997; 64: 21-30.
- Viteri G, Carrard G, Birlouez-Aragón I, Silva E, Friguet B. Age-dependent protein modifications and declining proteasome activity in the human lens. Arch Biochem Biophys. 2004; 427: 197-203.
- Stiuso P, Libondi T, Facchiano AM, Colicchio P, Ferranti P, Lilla S, et al. Alteration in the ubiquitin structure and function in the human lens: a possible mechanism of senile cataractogenesis. FEBS Lett. 2002; 531: 162-167.
- Babizhayev MA, Yegorov YE. Reactive oxygen species and the aging eye: specific role of metabolically active mitochondria in maintaining lens function and in the initiation of theoxidation-induced maturity onset cataract- a novel platform of mitochondria-targetedantioxidants with broad therapeutic potential for redox regulation and detoxification of oxidants in eye diseases. Am J Ther. 2016; 23: e98-e117.
- Ansari NH, Wang L, Srivastava SK. Role of lipid aldehydes in cataractogenesis: 4-hydroxynonenal-induced cataract. Biochem Mol Med. 1996; 58: 25-30.
- Sreelakshmi V, Abraham A. Cassia tora leaves modulates selenite cataract by enhancing antioxidant status and preventing cytoskeletal protein loss in lenses of Sprague Dawley rat pups. J Ethnopharmacol. 2016; 178: 137-143.
- Duncan G, Jacob TJ. Calcium and the physiology of cataract. Ciba Found Symp. 1984; 106: 132-152.
- Shukla N, Moitra JK, Trivedi RC. Determination of lead, zinc, potassium, calcium, copper and sodium in human cataract lenses. Sci Total Environ. 1996; 181: 161-165.
- Marcantonio JM, Duncan G, Rink H. Calcium-induced opacification and loss of protein in the organ-cultured bovine lens. Exp Eye Res. 1986; 42: 617-630.
- Sanderson J, Marcantonio JM, Duncan G. Calcium ionophore induced proteolysis and cataract: inhibition by cell permeable calpain antagonists. Biochem Biophys Res Commun. 1996; 218: 893-901.
- Duncan G, Wormstone IM. Calcium cell signalling and cataract: role of the endoplasmic reticulum. Eye (Lond). 1999; 13 : 480-483.
- Sreelakshmi V, Sasikala V, Abraham A. Luteolin supplementation prevents selenite- induced cataractogenesis in Sprague Dawley rat pups. ChemBiodivers. 2015; 12: 1881-1890.
- Zeng J, Zhang Z, Paterson CA, Ferguson-Yankey S, Yappert MC, Borchman D. Ca(2+)-ATPase activity and lens lipid composition in reconstituted systems. Exp Eye Res. 1999; 69: 323-330.
- Ahuja RP, Borchman D, Dean WL, Paterson CA, Zeng J, Zhang Z, et al. Effect of oxidation on Ca2+ -ATPase activity and membrane lipids in lens epithelial microsomes. Free Radic Biol Med. 1999; 27: 177-185.
- Cooper KE, Tang JM, Rae JL, Eisenberg RS. A cation channel in frog lens epithelia responsive to pressure and calcium. J Membr Biol. 1986; 93: 259-269.
- Barbusinski K. Fenton reaction-controversy concerning the chemistry. EcolChem Eng. 2009; 16: 347-358.
- Cook CS, McGahan MC. Copper concentration in cornea, iris, normal, and cataractous lenses and intraocular fluids of vertebrates. Curr Eye Res. 1986; 5: 69-76.
- Balaji M, Sasikala K, Ravindran T. Copper levels in human mixed, nuclear brunescence, and posterior subcapsular cataract. Br J Ophthalmol. 1992; 76: 668-669.
- Goralska M, Holley BL, McGahan MC. Identification of a mechanism by which lens epithelial cells limit accumulation of overexpressed ferritin H-chain. J Biol Chem. 2003; 278: 42920-42926.
- Goralska M, Nagar S, Fleisher LN, McGahan MC. Differential degradation of Ferritin H- and L-Chains: accumulation of L-Chain-Rich Ferritin in lens epithelial cells. Invest Ophthalmol Vis Sci. 2005; 46: 3521-3529.
- Er H, Gunduz A, TurkozY, Cigli, A, Isci N. Effects of NG-nitro L-arginine and corticosteroids on aqueous humor levels of nitric oxide and cytokines after cataract surgery. J Cataract Refract Surg. 1999; 25: 795-799.
- Horton JW. Free radicals and lipid peroxidation mediated injury in burn trauma: the role of antioxidant therapy. Toxicology. 2003; 189: 75-88.
- Ghafourifar P, Bringold U, Klein SD, Richter C. Mitochondrial nitric oxide synthase, oxidative stress and apoptosis. Biol Signals Recept. 2001; 10: 57-65.
- Ito Y, Nabekura T, Takeda M, Nakao M, Terao M, Hori R, et al. Nitric oxide participates in cataract development in selenite-treated rats. Curr Eye Res. 2001; 22: 215-220.
- Ghayur T, Banerjee S, Hugunin M, Butler D, Herzog L, Carter A, et al. Caspase-1 processes IFN-gamma-inducing factor and regulates LPS-induced IFN-gamma production. Nature. 1997; 386: 619-623.
- Gu Y, Kuida K, Tsutsui H, Ku G, Hsiao K, Fleming MA. et al. Activation of interferon-gamma inducing factor mediated by interleukin-1beta converting enzyme. Science. 1997; 275: 206-209.
- Lee EH, Wan XH, Song J, Kang JJ, Cho JW, Seo KY. et al. Lens epithelial cell death and reduction of anti-apoptotic protein Bcl-2 in human anterior polar cataracts. Mol Vis. 2002; 8, 235-240.
- Spector A. The lens and oxidative stress. Sies H, editors. In: Oxidative stress, oxidants and antioxidants. Academic Press, London, 1991; 529-558.
- Yan Q, Liu JP, Li DW. Apoptosis in lens development and pathology. Differentiation. 2006; 74: 195-211.
- Yao K, Tan J, Gu WZ, Ye PP, Wang KJ. Reactive oxygen species mediates the apoptosis induced by transforming growth factor beta(2) in human lens epithelial cells. Biochem Biophys Res Commun. 2007; 354: 278-283.
- Yao K, Ye PP, Zhang L, Tan J, Tang XL, Zhang YD. Epigallocatechingallate protects against oxidative stress induced mitochondria dependent apoptosis in human lens epithelial cells. Mol Vis. 2008; 14: 217-223.
- Arnold AC. Basic Principles of Ophthalmic Surgery. USA: American Academy of Ophthalmology. 2006.
- Singh SK, Winter I, Surin L. Phacoemulsification versus small incision cataract surgery (SICS): which one is a better surgical option for immature cataract in developing countries. Nep J Oph. 2009; 1: 95-100.
- Ruit S, Tabin G, Chang D, Bajracharya L, Kline DC, Richheimer W, et al. A prospective randomized clinical trial of phacoemulsification Vs manual sutureless small-incision extracapsular cataract surgery in Nepal. Am J Ophth. 2007; 143: 32-38.
- Chan E, Mahroo OA, Spalton DJ. Complications of cataract surgery. Clin Exp Optom. 2010; 93: 379-389.
- Melese M, Alemayehu W, Friedlander E, Courtright P. Indirect costs associated with accessing eye care services as a barrier to service use in Ethiopia. Trop Med Int Health. 2004; 9: 426-431.
- Weikel KA, Garber C, Baburins A, Taylor A. Nutritional modulation of cataract. Nutr Rev. 2014; 72: 30-47.
- Asha R, Devi VG, Abraham A. Lupeol, a pentacyclictriterpenoid isolated from Vernoniacinerea attenuate selenite induced cataract formation in Sprague Dawley rat pups. ChemBiol Interact. 2016; 245: 20-29.
- Sasikala V, Rooban BN, Sahasranamam V, Abraham A. Rutin ameliorates free radical mediated cataract by enhancing the chaperone activity of α-crystallin. Graefes Arch Clin Exp Ophthalmol. 2013; 251: 1747-1755.
- Manikandan R, Thiagarajan R, Beulaja S, Chindhu S, Mariammal K, Sudhandiran G, et al. Anti-cataractogenic effect of curcumin and aminoguanidine against selenium-induced oxidative stress in the eye lens of Wistar rat pups: An in vitro study using isolated lens. Chem Biol Interact. 2009; 181: 202-209.
- Doganay S, Borazan M, Iraz M, Cigremis Y. The effect of resveratrol in experimental cataract model formed by sodium selenite. Curr Eye Res. 2006; 31: 147-153.
- Rooban BN, Sasikala V, Sahasranamam V, Abraham A. Amelioration of selenite toxicity and cataractogenesis in cultured rat lenses by Vitex negundo. Graefes Arch Clin Exp Ophthalmol. 2011; 249: 685-692.
- Sasikala V, Rooban BN, Priya SG, Sahasranamam V, Abraham A. Moringaoleiferaprevents selenite-induced cataractogenesis in rat pups. J OculPharmacolTher. 2010; 26, 441- 447.
- Ertekin MV, Kocer I, Karslioglu I, Taysi S, Gepdiremen A, Sezen O, et al. Effects of oral Ginkgo biloba supplementation on cataract formation and oxidative stress occurring in lenses of rats exposed to total cranium radiotherapy. Jpn J Ophthalmol. 2004; 48: 499-502.
- Halder N, Joshi S, Gupta SK. Lens aldose reductase inhibiting potential of some indigenous plants. J Ethnopharmacol. 2003; 86: 113-116.