
Research Article
Austin J Clin Pathol. 2023; 10(1): 1079.
Future of AI: The Cell shows the Way
Ashok Kumar Mukhopadhyay¹*; Vivek Kumar²; Abha Singh³; Manish Ranjan³; Tapasyapreeti Mukhopadhyay¹; Abhijeet Kumar&³; Namrata Sarin³
¹All India Institute of Medical Science, New Delhi, India
²Indian Institute of Management, Jammu, India
³North DMC Medical College & HRH, Delhi, India
*Corresponding author: Ashok Kumar Mukhopadhyay All India Institute of Medical Science, New Delhi, India. Email: mukhoak1953@gmail.com
Received: June 05, 2023 Accepted: June 29, 2023 Published: July 06, 2023
Abstract
Systems cell is an easily available model to study the hierarchy and the detail in programming for Systems Governance. The idea of the system is usually conceived with a vision, and a “will” for showcasing it in the objective 4-D domain. When consciousness conceives the vision, its bosom-associate nascent nature inscribes the “will” into intent. Other associates prepare the logic and the logistics with consideration of ethics and aesthetics. The intent is then translated into informed instructions for briefing the members of different expert committees to achieve the desired goal of the system in their respective domain. The members meet, talk, exchange opinions, and prepare strategies for implementation of the vision. Following application of their mind, the information is converted into signals passed to the trained employees to conduct the eventful ceremonies almost effortlessly and in an automated manner. This model of programming is followed by a biological cell and the cell shows us the direction for the future of AI.
Keywords: Cell signaling; Organogram of organellosomes; Cell’s will; Autonomy; Holonomy; Systems Governance; Three-tier cell programing
Introduction
We begin with the research question, how the programming of a cell differs from a signal-based programmed machine available today? It is true that a biological cell is much more than an automated, signal-driven programmed machine! Cell’s intelligence is nature-made, and not artificially made by scientists! To achieve the intelligence of a biological cell, and intelligence in nature in an artificial device is a next-to-impossible project and would be a century-long journey! There are astonishing features in the behavior of even a single cell that point out that a cell can “will”, has choice, and makes decisions in complex situations. Pathology in this sense is not merely morbid biochemistry, physiology and anatomy! Pathology is the life story of cells, tissues, organs, and systems of the body [1]. In this paper, authors elaborate how the programming of a cell differs from a programmed signal-based machine placing some of the facts of cell biology and pathology into appropriate context. It is also proposed that the systems cell has an open-ended, three-tiered, and nested programming that can explain most of its behavior. The operation of each tier is conducted in a specific mode by a specific currency to achieve specific objectives.
Support from the Evidence Accumulated in Cell Biology
A cell obviously does not have a so-called ‘brain’ or brain-like structure. In spite of this, a cell is conscious, shows its will, has choice, can learn, makes decisions as evident from several published experimental works on the behavior of a simple unicellular slime mold, Physarum polyencephalum (brainless but multiheaded) [2,3]. A bacteria or protist can locate pray or potential mate, and has the ability to escape from predators [4]. Stentor Roseli exhibits complex avoidance behavior [5]. Even the slime mold has the ability to communicate with very long distant spatial information and generate similar patterns on its slime [6]. Habituation and sensitization do not require a cell to have neuron character [7,8]. This is observed in non-neural cells as well. In a tissue, organ and system, a cell can break its established pattern, and make a new pattern, which is termed differentiation.
The cell makes purposeful informative communication with its colleagues. It has been shown that the telomere is gifted by APC (Antigen presenting cell) to a T lymphocyte to keep the T cell young and the memory of contact with antigen longer [9]. Intercellular meaningful exchange happens through exosomes, e.g., Extracellular Vesicles (EV) from stem cell containing several species of RNAs (mRNAs, microRNAs and long non-coding RNAs) enter neighboring injured cells to reprogram it through epigenetic mechanism [10]. Exosomes of cancer cells and bone marrow-derived progenitor cells facilitate pre-metastatic niche formation and metastasis [11]. Senescent cell’s EVs promote senescence of other cells. Non-senescent cells’ EVs are seen to rejuvenate senescent cells [12]. The origin, structure and functions of EVs are described in the literature [13,14]. Neurons busy with ‘higher’ functions, get their major ATP supply free of cost from the astroglial cells. Beside Astrocyte-Neuron-Lactate Shuttle (ANLS) [15], astrocyte generates ATPs 20 times more than that produced by a neuron and they do it for sustaining their conjugate relationship for the sake of the optimal functioning of the nervous system they belong to. Where such higher functions are not needed and neurons transmit only signals, as in the ganglion (collection of nerve cells outside the nervous system), there is no need for such astrocytes around neurons! In necessity, cell to cell communication can reach such a level that even an important organelle such as the mitochondria could be transferred between metabolically rich and metabolically compromised cells [16]. Such intercellular mitochondria transfer is one of the mechanisms of immunometabolic crosstalk that is impaired in obesity.
One of the major scientific feats [17] in 2010 in the laboratory of J. Craig Venter Institute is the synthesis of the whole genome of Mycoplasma mycoides and the subsequent cloning of this DNA sequence inside the yeast cell and then transplanting the genome inside the Mycoplasma capricolum, whose own genome had already been removed. This new bacterial colony grew in culture. To clarify and emphasize, the feat requires an intermediate yeast cell. Also the growth in culture requires a mycoides carrier cell. This raises the issue of the difference between DNA as a chemical substance, and DNA in life-situations. The dilemma leads our imagination towards something subtle and intangible operating within an existing life-form. There are differences between the DNA as a chemical, and the DNA as an informational molecule (e.g., DNA buried in fossil), and the informative DNA molecule in life-situations. A chemical DNA can be made to replicate very fast almost endlessly in a PCR machine without error, but the same molecule of DNA, once put within protoplasm replicates very slowly in a limited way and that too not without flaws! A chemical DNA cannot be transcribed into mRNA and translated into a protein molecule without having the surrounding milieu of protoplasm, the principle used by Craig Venter to produce A-Life. Something subtle and intangible we are missing in the description of life merely as a life-form! The protoplasm is real, as well as vital in this context.
The cell, although, possesses several molecular robots and runs several signal-based programs by automated operations, has an extraordinary power of autonomy over such automation. The cell possesses signalosomes over the signal networks. Signalosomes consist of conformationally-equipped proteins that can possibly extract the meaning from a signal as a piece of information, and in reverse, probably can generate informative signals! There are several of such organellosomes (we are coining this new term in cell biology) floating within the protoplasm of the cell such as nucleosome, centrosome, ribosomes, proteasomes, signalosome, peroxisome, lysosome, inflammasome etc. They operate as “perceiver”, in contrast to several proteins which have been identified to act as sensors.
There might be a debate whether a signal-driven, neural network-based programmed machine “learns” by “perception” of the environment which is popularly known as machine-learning, or, is it a kind of passive familiarization of a signal-recognizing-operative device with different prospective signal patterns? Is there a mechanism of building up any memory, and its retrieval in such a device? According to Edo Liberty, the founder and CEO of Pinecone, “While AI models such as GPT from Open AI are trained on billions of pieces of data, they don't remember anything you show them or even anything they give back to you. AI models are stateless. They have no memory.” (20th Mar-2023). On the other hand, in the live-situations the debate continues whether a cell distinguishes “self” from “non-self” on the basis of conscious perception, and actively acquired memory and experience, or by passive recognition of mere molecular patterns namely Damage Associated Molecular Pattern (DAMP) and pathogen associated molecular pattern (PAMP)? There exists a mechanism of building up a long term memory (e.g., in memory T cell), prospective memory and its retrieval in the cellular systems.
Leaving aside the debates, the prudent direction of investigation would be to raise research questions on the difference between signal and information, between a sensor and a perceiver, and when and how a non-informational molecule of the cell becomes an informational molecule, a sensor protein becomes a perceiver protein? What conformational change in the molecule brings such a change! When does the postulated “conformon” [18] of Ilya Prigogine appear in the cell-scenario [19,20,21]? What finances this operation for conformational change?
An automated programmed robot does not go through the process of survival and death. It is extremely stable as compared to a cell! In some sense, this might be an advantageous position for a robot! A cell, on the other hand, with so many molecular robots operating within it, is always hanging in the balance of survival and death! The cell has to be engaged incessantly in uncertainty homeostasis for survival. There are injurious symmetry-breaking processes, and the cell makes new symmetry for homeostasis. Such homeostasis failure leads to neurodegeneration and even malignancy! There are disorders of cellular autonomy which when incorrigible proves lethal to the system.
Support from the Alternative Interpretation of Pathological Processes
The autonomy of a cell works, however, within the holonomy of the tissue, organs, and the systems of the body. An autonomous cell does not encroach upon the autonomy of other members of its tissue, organ or system. When it does so, it creates an example of autocracy manifested as dysplasia, and even what is called malignancy. There are molecular check points on the route of this happening, executed by the gatekeeper gene (e.g., APC gene), which stops cell’s G0>G1 transition, the guardian gene (e.g., P53 gene), which recommends repairing of misdirected “damaged” DNA and if that too fails directs the cell for apoptosis. Lastly, there is the Governor gene (e.g., Rb gene), which finally restricts cells not to continue with the cell cycle in G1>S restriction point if their DNA is unrepairable. or in G2>M restriction point when their DNA remains unduplicated or damaged in S phase of cell cycle. If somehow, the cell’s autocracy overcomes all check points, the outcome is what we call a malignant cell. We have some understanding in biology on how autonomy transits to autocracy. For future perspectives, our research question is how a cell accommodates a congregation of so many automated operations flawlessly, and maintains its own autonomy over several automated molecular robots and their operations? Generating evidence for answering such a question would be a new frontier in cell biology!
As stated, sensation and perception are different. Sensation is signal-driven. Perception is information-driven. Learning from sensations and learning by perception are distantly different. Sensation-based learning could be best called training. Perception-based learning creates memory. Building of memory is a part of education! Without memory there is little learning or any education! Fundamentals can be learnt by perception, not by sensation. Muscle contraction as a result of nerve conduction is an example of signal transmission. The reflex development of such activity is the core part of any training. Perception, on the other hand, is a ‘brain’-phenomenon, and on a finer note, is a psychic phenomenon. A lizard camouflages itself differently when it encounters a prey, an enemy, and a sex-mate. This is the result of learning by perception. Sensation can be mechanized by placing sensors in appropriate positions. Perception is a biological property derived from consciousness, and requires a perceiver. Physiologically, sensation is the outcome of signal processing while for perception one requires information processing. In cell biology one can distinguish signal molecules (e.g., a peptide chain) from informational molecules (e.g., folded protein). The switch-over from the discipline of biochemistry to the discipline of molecular biology happens exactly on this crucial point. Biochemistry deals with non-informational molecules, while molecular biology deals with informational molecules only.
To err is a property of an automated machine! It is a passive phenomenon! To make a mistake (omission?), or blunder (commission?) is an active and conscious-centered phenomenon characteristic of a conscious entity, and so of any biological cell. While a machine, such as a blood cell counter or a biochemistry autoanalyzer makes only errors in a random (random variability) or systematic (bias) way, a living entity during decision-making and subsequent behavioral expression, often commits a mistake or even a blunder. An automated signal-based machine does not have the power of perception and so it is in an advantageous position not to make any mistake or blunder. Which can never happen in a machine is to perceive a friend as an enemy, a mistake, or to perceive an enemy as a friend, a blunder. Pathological processes might originate from the faults in perception of its surrounding environment by the cell. In immunobiology, mistaken perception of a friend as enemy is observed in the pathogenesis of autoimmunity, lymphocytes recognizing the “non-self” patterns as the patterns of “self”! At the next control level, the regulatory T cell presses the accelerator instead of pressing the brake! Autoimmune damages start! A blunder of perceiving an enemy as a friend is observable during the immune-bypass mechanism of malignant cells. At the regulatory T cell level where the accelerator was supposed to be pressed to destroy the malignant cells, the brake has been pressed! The result of this blunder is obvious, the enemy captures the scenario!
A complexly developed automated robot cannot engage in any kind of homeostasis with the environment at the level of deep physics. The present day robots never can participate in uncertainty-certainty homeostasis, symmetry-breaking and symmetry-making homeostasis, and intangible-tangible energy homeostasis. The robot does not have any access to intangible dark energy! Therefore, a robot never suffers from anxiety, stress or depression. Human beings suffer! A cell suffers. For survival, and to avoid death the cell is continuously engaged in the above-mentioned three homeostasis, the failure of which pushes the cell into the clinic/ward of cellular emergency medicine (G0 phase?). Anxiety is the result of homeostatic failure in the context of uncertainty-certainty. Stress arises with the failure of symmetry homeostasis that leads to pathological conditions like neurodegeneration, inflammatory bowel disease, infection like tuberculosis, and even malignancy. Depression originates from the repulsive property of accumulated dark intangible energy that cuts off the ‘self’ from environmental signals, sensation, and tangible energy. Such a depressive state, in serious situations, pushes out the enzyme cytochrome C from the space within mitochondrial double membrane into cytosol, which pushes the cell towards suicide, we mean apoptosis. Anxiety, Stress and Depression are thus whole body disease, which initially might begin with the neurons in the brain [22].
An automated intact machine in operative state does not have access to Zero-Point Energy (ZPE). An intact functioning biological cell has! That is why the cell can recover from broken symmetry, various imbalances of tangible energy and transform some uncertainties into certainty! For an artificial model of intelligent automation, the sensible signal-based material world stops at zero-point energy, at the cosmological constant of Einstein! This could be described as RIP (rest in peace) state for the signal-based material machines! The situation is certainly not the same for the cellular molecular machines whose operations are not merely signal-based, but also information-based. The cell might go back to its cellular zero-point energy state for taking rest, and its molecular machines continue to operate. Zero-point energy is supposed to be the door of communication between systems biology and systems cosmology, and systems physics and systems psychology [23].
A machine, automated and flawless, does not have any feeling and, therefore, cannot express emotion! On the other hand, the cell, for example a macrophage, often shows emotional expression. A macrophage shows its emotional frustration when it cannot engulf or chew a foreign material. The pathogenetic mechanism of almost all occupational lung diseases, especially asbestosis is this “frustrated” phagocytosis when there is outpouring of various lethal enzymes from the macrophage which initiates local pulmonary tissue damage and inflammation [24]. A normal monocytic cell is emotionally nonviolent. The monocyte may become violent in certain stimulated states as happens in some cases of COVID-19 infection [25], when a macrophage indiscriminately phagocytosed RBCs and other WBCs creating Hemophagocytic Lymphohistiocytosis (HLS), and induced a cytokine storm.
The Proposition
Automation, autonomy and holonomy are nested three goals in the programming of any advanced intelligent device. The programming of an automated machine is signal-driven, run according to the algorithm constructed by logic under the guidance from a neural network model of Governance. The purpose is to make the operation or constellations of operations effort-free and error-free, as much as possible, with a high yield per unit of time. The logic, however, is defunct without a supporting medium. With the materialistic base of such programming, the medium is the so-called celebrated “ether”, a better name would be etheroplasm! The description completes the programming in the nest I. This feat is almost achieved in the artificial devices of intelligence as available today.
The goal in the tier II programming is to gain autonomy over multiple interconnected automated operations. To begin the program, the system needs to understand the meaning of the operations going on. This itself is a great feat! It calls for some kind of awareness of the system itself and the environment. What it essentially requires is the supporting medium of protoplasm of a living cell. The operations are run by available logistic rather than algorithmic logic, both are derivatives of the sense of ethics and aesthetics originating from the nest III. The logistics followed are inclusive of ethics and aesthetics. The currency of the operation shifts from the signal to information. The device is governed by the model of an organogram. The purpose is to achieve systems perfection in terms of minimization of the probabilities of mistake and blunder, contextually assessed by locally relevant ethics and aesthetics. To have this protoplasm as a medium appears to be the first step in technology of a science for consciousness, which the human being is yet to take. Without such a medium of protoplasm, the organogram is defunct, the logistic is empty and the currency of information is nonfunctional. All fulfilled, we are towards developing a model of cellular intelligence.
In tier III programming, the goal shifts from autonomy to holonomy. Holonomy here is meant as autonomy within the systems whole that makes the systems sustainable with multiple autonomous orders. In this sense, holonomy is respecting each other’s autonomy in composition of several autonomous components of the whole. The currency of tier III programming is intention scaled out of the ‘will’. This intention carries the ultimate wisdom and the purpose of the system. Three together percolates as the “intent” in the currency of information in tier II. Tier III is operated by three operators, the sentient-entity, philosophically named “self”, the homeostatic entity, scientifically called “life”, and the event-making entity, popularly known as “mind”. The Governing authority of tier III is consciousness. The supporting medium is “psychoplasm”, a subtler form of the cellular protoplasm transited through ZPE. The operators and the operations are truly nonlocal within the systems cell, systems being, as well as in the system-independent domain of the world. As big is the “whole”, so big is the ambition to achieve success in this programming!
Intellectually comprehensible largest system, as known, is the systems of multiple universe(s). In this construction, the traceability of the psychoplasm is with the source, the multiversal plasm, which the primary author of this paper has named the essence of the Multiversity in 1995 [26]. The postulated supporting background of the material world is ether (etheroplasm). The real background of life-form is protoplasm. The traceability of all psychoplasm, protoplasm, and etheroplasm is with the Essence of the Multiversity through ZPE (Figure 1).
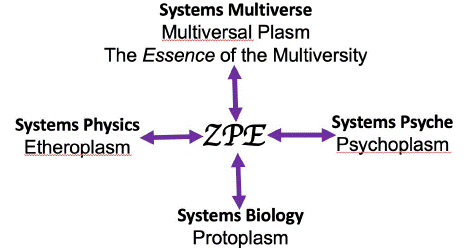
Figure 1: Since “life” is naturally outside the space time world, a biological cell has natural access to Zero-Point Energy state (ZPE). At ZPE, Systems Physics meets Systems Psyche, and Systems Biology meets Systems Cosmology. Here, we find the traceability of Etheroplasm of the material world, Protoplasm of the biological world, and the Psychoplasm of the psychic world into the Multiversal plasm of the cosmic world.
The intelligence in tier III could be called holonomic natural intelligence! The memory is holographic [27]. What has been said in this proposition has been summarized in Table 1. Tier I is the most superficial nest, and tier III is the deepest nest. Sandwiched in between is the nest of tier II.
![]()
TIER - I
TIER - II
TIER - III
The Goal
To achieve effortless, automated and error-free results with maximum yield
To achieve Autonomy over automation
To achieve sustainability, harmony with other autonomous components; Holonomy
Operations run on the basis of
Sensation,
LogicPerception,
Logistic based on the Concept, Hypothesis, and Theory of the System.“Will” based on the Wisdom of the system and “Intention” that carries the purpose
Supporting Medium
Etheroplasm
Protoplasm
Psychoplasm
Currency
Signal
Information
Intention
Governance
Neural Network Model of Governance
Organogram Model of Governance
Operative model of Non-Observable but Influential Actors
Control
Feedback control of signaling
In addition to feedback, there are feed forward controls.
Protoplasm is involved in the control processHolographic control
Operators in biological cell
Polypeptides and Receptor proteins etc.
The orchestra of organellosomes
Consciousness, Self, Life, and Mind
Mode of Communication
Signal networking
‘Molecular Talk’ is a misnomer phrase
“Personalized” reach to others for ‘talking, to share information. “Organellosomes talk”,
“Cellular talk”Contagiousness
of IntentionalityIntelligence
Model of Intelligence in Artificial device
Cellular or Biological Intelligence
Natural Intelligence
Room for Emotion
Nil
Present
Apparently not.
At deeper level, fine-tuned Emotion!Nature of Memory
Working memory/sensory memory.
Measured by byte.
Explicit memory.Memory bridging the past with the present
Implicit memory.
Semantic/Informational memory.
Episodic/experiential memory.
Prospective memory through long term biological membrane potentiation bridging the present with the futureHolographic memory.
No need of retrieval.
It is thereIn terms of depth of Physics
Physical (Classical & Quantum) nests of nature
Subquantum/Pre-quantum nest of nature
Sub-subquantum nest of nature
Table 1: Nested, Three-Tiered, Open Programming as Observed in a Live-system.
Explaining the Nests in a Biological Cell
Characteristically and exemplarily the cell is programmed in a nested three tier way. Cell’s cognitive functions such as will, intention, choice, decision etc. are programmed in tier III. Cell’s informative activities are programmed in tier II, operated by the orchestra of organellosomes. Cell’s automated functions are conducted in tier I through molecular robots by the currency of signal-, and receptor-molecules, mostly peptides and folded proteins respectively. The pathological processes happening in tier I are corrigible by tier II activities while the tier II problems are fixed from the operators in tier III. The issues in nest III get sorted out spontaneously to some extent due to the system’s openness to the nonlocal world (Figure 2). Let us describe the programming in detail.
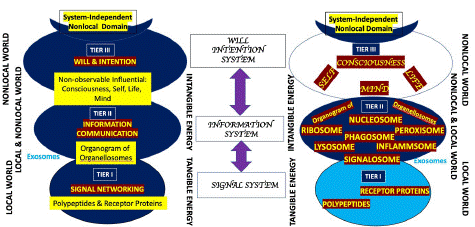
Figure 2: The programming of a cell is open-ended, three tiered, and nested. As shown in the figure, nest II is larger than nest I and nest III is larger than nest II. Nest III is open to the system-independent nonlocal world. Tier one is a signal-driven world. Nest I in biology is run by signal and receptor molecules. Nest II is information-driven, and in a biological cell is run by the organogram of the organellosomes. Tier III is a miniature form of human psyche with non-observable but influential operators like Mind, Self, Life and Consciousness. This nest is will and intention-driven. Over the spectrum as one goes from superficial to deeper, a transition is observed from tangible to intangible energy, local domain to nonlocal domain.
Tier III: The deepest and the third tier of cellular programming is run by the currency of ‘intention’ and ‘will’ of the non-observable but influential operators and their operations. Mostly these are cognitive functions, and require participation of the event-making entity, mind, to connect with information at the final common path to subsequent nests. The operating members of tier three are nonlocal, meaning they cannot be localized in terms of space and time. They maintain the connection and communication of a living entity with the nonlocal world. An entity which acts locally but has the ability to communicate nonlocally reserves the capacity to evolve: the most secret power of evolution is thus demystified. It also supports Einstein’s statement, “The true sign of intelligence is not knowledge but imagination.” The insights from the already mentioned recent advances in cellular behavior [2-16] lead us to formulate a research hypothesis that the apparently non-observable but influential operators and the operations which are understood to use the brain of a human being to execute a behavior [28,29] can also use a single cell to ensure a similar behavioral output in a small cellular scale. Probably this is because the constitution of the behavior of the being is written in the constitution of its constituent cells in a holographic way. Nest III defines the frontier in psychology.
Tier II: The second tier of the cell is characteristically governed by the organogram made of the members of the ‘oganellosomes’ of the cell that operate by contact and “talk” through the currency exchange of information. They respond by moving at the spot, when and as necessary. Ribosomal RNAs move from nucleolus to ribosomes in the cytoplasm. Ribosomes move along the endoplasmic reticulum. Phagosome moves to lysosomes or vice versa for fusion!
Probably, all such ‘-osomes’, form an orchestra of the oganellosomes carrying out their activities through contact and information exchange by personalized “talk”. The mode of communication of the members of the organellosomes might be cited as an example of cell-mimesis, which in the case of non-movable neurons is close to neuromimesis [30]. Each organellosome is rich in quaternary-structured/spherical protein, and is suggested to have the operative capacity to extract meaning out of signals! The combination inside a ‘-osome’ operates as a perceiver in contrast to a sensor. The combinations act as informational nodal points within the protoplasm, in contrast to nodal points in neural network model operating for signals. Such spotlights within an informational network are seen as orchestration of nucleosomes [31] floating within nuclear sap, and creation of centrosomes during mitosis. Proteasomes are available both within the nucleus and cytoplasm. Inside the cytoplasm, the orchestra is constituted by proteasomes, ribosomes [32,33], peroxisomes [34], signalosomes [35,36], phagosome–lysosomes [37], inflammasomes [38], etc. The role of exosomes as EV has already been described [10-12]. Microsomes, liposomes and synaptosomes are experimental creations of such “-osomes” to study the membrane function alteration, lipid handling, and synaptic activities respectively. Physically, all of the organellosomes have spherical or spheroidal shape. Chemically, they consist of highly folded, meaning ‘experienced’ proteins. All of them have a surrounding membrane, except ribosomes and proteasomes, both move around the membranous structures. While error happens during automated signaling, the mistakes and blunders in perception are suggested to happen at this second tier of organogram of such organellosomes. It seems that the organellosomes are devised for achieving ‘autonomy over the automation’! Nest II determines the frontiers in cell biology, especially relevant in modelling of intelligence.
Tier I: At present, the major point of attraction of cell biologists is the most superficial nest of programming of the cell. Cell signaling, signal transduction, signal interception, and the sensor proteins are prime topics of research. The communication between the receptors is almost similar to the signal networks of automated machines, except the presence of supporting medium of protoplasm. Cell signaling is mostly executed by polypeptide molecules. For generation of information from any signal, the milieu of protoplasm is essential along with informational nodes such as signalosomes where the signal could be read by a cell as information.
Perspectives
Intelligence as such is difficult to observe in an artificial device. What is seen is the intelligence of the programmer in the program. Intelligence is obviously biological and also natural. Artificial Intelligence (AI) is a “marketing term” (Federal Trade Commission, USA, 2023). In this context, the Cellular Model of Programming (CMP) offers us the direction for the future of the Artificial Devices of Intelligence (ADI). This paper could be further developed on the basis of practical distinctions drawn between signal and information, sensation and perception, a sensor and a perceiver, error and mistake, life-form and ‘life” in subtle phase, logic and logistic that led the author to a position to distinguish an Artificial Device of Intelligence (ADI) from the real Biological and Natural Intelligence (BI and NI respectively). With the proposition of the functioning of organellosomes as an organogram, this paper threads the entire community of cell organelles into an orchestra. In the process, it opens up new frontiers in cell biology, psychology and synthetic biology. The creation of protoplasm has been recognized as the first step forward for synthetic biology which is essential for stepping up to gain autonomy over automation. An important feat in technology of the synthetic biology frontier would be to see whether the organellosomes recovered from a cell can be made to work without the medium of protoplasm!
We may investigate whether any synthetic plasm manufactured for this purpose can have an access to ZPE or could it be churned and modified by directed neutrino beams? Access to ZPE and getting influenced by natural neutrino showers are deep properties of a cell. Made possible in the artificial devices of intelligence solves several problems by mechanizing the homeostasis of uncertainty-certainty, symmetry-breaking and making, and the economics of intangible and tangible energy.
We are standing on the truth of this great insight; life-form, although, is bound by space and time, the “life” itself, in its subtler phase, is naturally outside the space-time world. For the same reason, the live entities, when required, in an intact operative state can instantaneously contact zero-point energy, which a non-living machine cannot! This access is easier for a single cell than a compound tissue, or a system! That explains why Albert Einstein, a celebrated scientist of this community of the living beings, can draw the entire canvas of space-time-matter-energy science and discover cosmological constant (ZPE) objectively in the third person’s perspective with little trace of subjectivity.
In contrast to the widely presumed idea, DNA is not life! DNA is, however, that substance, which is essential for the fusion of the material and protoplasmic worlds! The spherical histone protein, the embodied wisdom (most compact crystallized information manifold), is the trusted companion of DNA in its journey into the protoplasm of life. In this perspective, the production of artificial model of Life and Artificial model of Intelligence is fused, and is an extremely ambitious project expected to succeed over a span of one century! Prior to this happening, the “Psyche” will be “out” there in this twenty-first century, with the objectivity in the third person’s perspective. Psychology then will no longer be labelled as a stream of humanities, but will be generously considered as a discipline of science intertwined with the science of information!
Mind is the final common path for the psyche, which has been gradually becoming “out” there, with application of several neurotechnologies, such as mind-reading machine, brain-machine interface, neurolinks, and chips in the brain. Many of those are found useful in management of Autism, Depression, Dementia, and even Obesity! Even in such a situation, human consciousness will remain as the last fortress of privacy affirming forever the human right to cognitive liberty.
Mind is not confined to neural ensembles called brain [7,8,39]. Every cell has its own mind and a language. It manages all its activities along the cognitive ladder of language, starting bottom-up from a Signal (space-time construct of information, digitized information), climbing to Information (non-digitized information that is factorizable into its three folia; content, intent and the ability to reduce uncertainty), Knowledge (symmetry of interrelated non-digitized and non-factorizable information), Experience (Information manifolds), and finally, the Wisdom (information crystal) [40,41]. The Top-down descent of the pattern has also been made possible in a cell. Unicellular slime mold can produce patterns of distant space in its slime [6]. However, even such a compact system like a cell can fall prey of plagiarism, e.g., in case of viral infections when several cell machineries speak the viral language. How the cellular language system could be of help for Large Language Model (LLM) of the Artificial Device of Intelligence (ADI) throws great challenges to the scientists. Our goal is not to manufacture a humanoid, but to humanize the ADI. The cell shows the way.
Author Statements
Funding & Conflict of Interest
No Funding Agency has funded this work. There is no conflict of interest between the authors in publishing this Article. Dr. Ashok Kumar Mukhopadhyay conceived the idea, composed the draft and improved it to the final stage of publication with constant constructive criticism of the co-authors.
References
- Mukhopadhyay AK. Humanities and spirit in cell science. A cell could be considered a universe for learning behavior. CPQ Neurol Psychol. 2022; 5: 01-23.
- Nakagaki T, Yamada H, Tóth A. Maize-solving by an amoeboid organism. Nature. 2000; 407: 470.
- Awad A, Pang W, Lusseau D, Coghill GM. A survey on Physarum polycephalum intelligent foraging behaviour and bio-inspired applications. Artif Intell Rev. 2023; 56: 1-26.
- Tang SKY, Marshall WF. Cell learning. Curr Biol. 2018; 28: R1180-4.
- Dexter JP, Prabakaran S, Gunawardena JA. A Complex hierarchy of avoidance behaviors in a single-cell eukaryote. Curr Biol. 2019; 29: 4323-4329.e2.
- Schenz D, Nishigami Y, Sato K, Nakagaki T. Uni-cellular integration of complex spatial information in slime moulds and ciliates. Curr Opin Genet Dev. 2019; 57: 78-83.
- Eisenstein EM, Brunder DG, Blair HJ. Habituation and sensitization in an aneural cell: some comparative and theoretical considerations. Neurosci Biobehav Rev. 1982; 6: 183-94.
- McFadden PN, Koshland DE Jr. Habituation in the single cell: diminished secretion of norepinephrine with repetitive depolarization of PC12 cells. Proc Natl Acad Sci USA. 1990; 87: 2031-5.
- Lanna A, Vaz B, D’Ambra C, Valvo S, Vuotto C, Chiurchiù V, et al. An intercellular transfer of telomeres rescues T cells from senescence and promotes long-term immunological memory. Nat Cell Biol. 2022; 24: 1461-74.
- Camussi G, Deregibus MC, Quesenberry PJ. Role of stem cell-derived extracellular RNA-carrying vesicles in cell reprogramming. Austin J Clin Pathol. 2014; 1: 1001.
- Peinado H, Lavotshkin S, Lyden D. The secreted factors responsible for pre-metastatic niche formation: old sayings and new thoughts. Semin Cancer Biol. 2011; 21: 139-46.
- Oh C, Koh D, Jeon HB, Kim KM. The role of extracellular vesicles in senescence. Mol Cells. 2022; 45: 603-9.
- Doyle LM, Wang MZ. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells. 2019; 8: 727.
- Borges FT, Reis LA, Schor N. Extracellular vesicles: structure, function, and potential clinical uses in renal diseases. Braz J Med Biol Res. 2013; 46: 824-30.
- Genc S, Kurnaz IA, Ozilgen M. Astrocyte – neuron lactate shuttle may boost more ATP supply to the neuron under hypoxic conditions – in silico study supported by in vitro expression data. BMC Syst Biol. 2011; 5: 162.
- Brestoff JR, Wilen CB, Moley JR, Li Y, Zou W, et al. Intercellular mitochondria transfer to macrophages regulates white adipose tissue homeostasis and is impaired in obesity. Cell Metab. 2021; 33: 270-282.e8.
- Gibson DG, Glass JI, Lartigue C, Noskov VN, Chuang RY, et al. Creation of a bacterial cell controlled by a chemically synthesized genome. Science. 2010; 329: 52-6.
- Volkenstein MV. The conformon. J Theor Biol. 1972; 34: 193-5.
- Ji S. The conformon as the atom of life: a hypothesis. Scientific Publishing Basis of Consciousness Forum; 2023. Google. Available from: https://mail.google.com/mail/u/0/?tab=rm&ogbl#search/sungchul/FMfcgzGsmWwpBpLzWWRlwVpJmtXTJVSJ.
- Ji S. The cell language theory. London: World Scientific Publishing Europe Ltd. 2018.
- Ji S. Free energy and Information Contents of Conformons in proteins and DNA. Biosystems. 2000; 54: 107-30.
- Mukhopadhyay AK, Ranjan M, Kumar A, Singh A, Mukhopadhyay A. Physics life psychiatry: cellular model and the ZPE. Arch Neurol Neurosci. ANN MS. 2023; 15: ID.000860.
- Mukhopadhyay AK. Einstein’s abandoned otherworld: how conscious ”will” manifests in a 4-D world?. EC Psychol Psychiatry. 2022; 11: 01-8.
- Mularski A, Marie-Anaïs F, Mazzolini J, Niedergang F. Observing frustrated phagocytosis and Phagosome Formation and closure using total internal reflection fluorescence microscopy (TIRFM). Methods Mol Biol. 2018; 1784: 165-75.
- Soy M, Atagündüz P, Atagündüz I, Sucak GT. Hemophagocytic lymphohistiocytosis: a review inspired by the COVID-19 pandemic. Rheumatol Int. 2021; 41: 7-18.
- Mukhopadhyay AK. Conquering the brain. Towards the essence of the multiversity and the Akhanda paradigm. New Delhi: Conscious Publications; 1995.
- Mukhopadhyay AK. Information holograph. The Structure, the source and its operation. A international journal of Bio Engineering, Neuro Sciences and technology. 2012; 2: 12-32.
- Mukhopdhyay AK. Consciousness cognition and behavior. Consciousness, cognition and behavior. J Neurosurg Imaging Tech. 2020; 6: 302-23.
- Mukhopdhyay AK. Cognitive canvas: molecular embroidery, fabric and the base. EC Psychol Psychiatry. 2018; 7: 428-39.
- Reilly C. Neuromimesis: picturing the humanities. Picturing the brain. Front Integr Neurosci. 2022; 16: 760785.
- McGinty RK, Tan S. Nucleosome structure and function. Chem Rev. 2015; 115: 2255-73.
- Lafontaine DLJ, Tollervey D. The function and synthesis of ribosomes. Nat Rev Mol Cell Biol. 2001; 2: 514-20.
- Hurt E, Cheng J, Baβler J, Iwasa J, Beckmann R. Eukaryotic ribosome biogenesis. Cell. 2023; 186: 2282-2282.e1.
- Mast FD, Rachubinski RA, Aitchison JD. Peroxisome prognostications: exploring the birth, life, and death of an organelle. J Cell Biol. 2020; 219: e201912100.
- Ma Y, He K, Berkowitz GA. Editorial: From structure to Signalsomes: new perspectives about membrane receptors and channels. Front Plant Sci. 2019; 10: 682.
- Kandy SK, Janmey PA, Radhakrishnan R. Membrane signalosome: where biophysics meets systems biology. Curr Opin Syst Biol. 2021; 25: 34-41.
- Nguyen JA, Yates RM. Better together: current insights into phagosome-lysosome fusion. Front Immunol. 2021; 12: 636078.
- Tweedell RE, Kanneganti TD. Advances in inflammasome research: recent breakthroughs and future hurdles. Trends Mol Med. 2020; 26: 969-71.
- Ciaunica A, Shmeleva EV, Levin M. The brain is not mental! coupling neuronal and immune cellular processing in human organisms. Front Integr Neurosci. 2023; 17: 1057622.
- Mukhopdhyay AK. Science of information. Global Journal of Science Frontier Research: A Physics and Space Science. version 1. 2021; 21: 15-30.
- Mukhopadhyay AK, Mukhopadhyay AS. Visualizing information as a dynamic entity road map of deep science, AI and humanity. Psychol Behav Sci Int J. 2019; 13: 555867.