
Review Article
Austin J Orthopade & Rheumatol. 2014;1(2): 5.
Strength versus Endurance Training
Ahmad S1* and Roach R1
1Department of Trauma and Orthopaedic Surgery, Princess Royal Hospital, United Kingdom
*Corresponding author: Ahmad S, Department of Trauma and Orthopaedic Surgery, Princess Royal Hospital, Telford, Shropshire, TF1 6TF, United Kingdom
Received: October 20, 2014; Accepted: November 26, 2014; Published: December 02, 2014
Abstract
The molecular level research is supporting the macroscopic and microscopic findings on whether strength training and endurance training are mutually exclusive. Certainly there has been much progress since the publication of Dudley’s (Sports Med 1987;4:79-85) work. The impression is that there are both central and peripheral factors. At the central level evidence is relatively limited but does seem specific. However, in the neuromuscular setting there are multiple interactions at a variety of levels involving at least three different pathways in muscles and excitatory/inhibitory links all the way to the promotion of mRNA activity. All these steps therefore go against strength and endurance training adaptations being mutually exclusive. If we however just focus on the single-fibre microscopic end result, changes do indeed appear very specific to the demands placed on the body. This fits with the adaptive plasticity of many systems during and after development. This is then affected by additional variables to give the macroscopic changes we have known about for some time. A very complex continuum of load versus response therefore exists with multiple controlling variables. The idea of a ‘single’ switch remains elusive.
Keywords: Human training; Macroscopic; Microscopic; Molecular
Introduction
The dilemma as to whether strength training and endurance training are mutually exclusive has been topical for the last three decades and indeed has generated much research that was initially observational and is now very much immunohistochemical. As a result researchers are moving from the field of applied physiology through to molecular biochemistry and genetics.
In order to answer the question posed we will briefly describe both types of training and their relevance before splitting the evidence into three stages, which to a certain extent reflects historical progress namely macroscopic evidence, microscopic evidence and molecular research.
Strength and Endurance Training
Strength training has the intention of getting stronger by generating more force across a joint to move or resist an object that can be directly measured. It is agreed that to undergo adaptive change the musculoskeletal system has to be regularly overloaded safely. Training is therefore explosive and immediate, hence anaerobic, against either weights or resistance. Typically this involves low repetitions of high force on a programme of a few (or even one) cycles to ensure adequate recovery that is tailored to the individual (periodisation). The end result is the typical physic of a body builder. For instance, Arnold Schwarzenegger’s competition body measurements were as follows: weight 106.5 kg, chest 145 cm, arms 56 cm, waist 84 cm, thighs 72 cm and calves 51 cm. These strength individuals tend to have a high percentage of fast twitch (type IIa and IIb)) muscle fibres (Table 1). High intensity strength training can however result in cardio respiratory responses [1].
![]()
Type I
Type IIa
Type IIb
Motor Unit Size
Small
Larger
Largest
Mitochondria Density
High
Low
Low
Fatigability
Fatigue-resistant
Fatigable
Fatigable
Aerobic Capacity
High
Low
Lowest
Anaerobic Capacity
Low
Medium
High
Contraction Speed
Slow
Fast
Fast
Contraction Strength
Low
High
High
Table 1: Skeletal Muscle Fibre Types.
Endurance training is seen as being at the other end of the spectrum of performance namely the athlete who can perform for hours with the appearance of an ultra-distance runner. The individual therefore must have adaptations to remain aerobic. Training is usually numerous repetitions of low intensity and lots of cycles. Endurance athletes possess a higher proportion of slow twitch (Type I) muscle fibres.
A requirement between these two extremes is usually more common as sports dictate a combination of these. For example the footballer or triathletes who have to balance levels of these extremes and be specific to the upper or lower limbs accordingly. Moreover, a sport can have individuals from one end of the spectrum to the other in 2 disciplines, such as road cycling (Bradley Wiggins) and track cycling (Sir Chris Hoy). Variances in appearance do exist however in every sport, for example Usain Bolt versus his Jamaican team-mates and in the Olympic velodrome sprint final in 2012 between Jason Kenny and Gregory Bauge. Cross training is present in all routines and explains why so much interest has been placed on this topic and why coaches keep elite training schedules a closely guarded secret.
Macroscopic
Macroscopic evidence refers to end point studies determining the final result of adaptations that occur with specific training modalities. This helped us initially to develop the ideas behind strength training and endurance work and the relevance of concurrent training and background physiology to anaerobic and aerobic exercise.
Typical macroscopic findings to endurance training include a reduction in heart rate, increased stroke volume and cardiac output, polycythaemia and a reduction in total body fat. Changes are widespread and can be split into both central and peripheral [2,3]. The marathon runner is the characteristic endurance athlete and these individuals have a low body mass index (BMI).
Strength training, however, uses body building as the ultimate example does not result in these adaptive changes, with the individual predominantly having muscle hypertrophy and more peripheral changes [2,3]. It is not just in weight lifters that these typical appearances are seen though; take 6-time track cycling Olympic gold medallist Sir Chris Hoy whose thigh size is 69 cm and is able to leg press a remarkable 631 kg.
VO2 max levels are another macroscopic end result of training. It has been defined as being the maximal rate of oxygen consumption during incremental exercise and is considered to be a reflection of the aerobic fitness of an individual. Wide variations are seen in VO2 max between sports. Generally, an increase in VO2 max is seen in endurance sports, whereas strength training has little effect on VO2 max. Table 2 shows the difference in VO2 max between athletes and non-athletes, males and females, and endurance and strength sports.
![]()
Sport
Age
Males
Females
Non-Athletes
10-19
47-56
38-46
20-29
43-52
33-42
30-39
39-48
30-38
40-49
36-44
26-35
50-59
34-41
24-33
60-69
31-38
22-30
70-79
28-35
20-27
Athletes (Endurance)
Running
18-39
60-85
50-75
Cycling
18-26
62-74
47-57
Skiing
20-28
65-94
60-75
Athletes (Strength)
Weightlifting
20-30
38-52
_
American Football
20-36
42-60
_
Discus
22-30
42-55
_
Shot Put
22-30
40-46
_
Table 2: VO2 max (ml/kg/min) in various individuals.
Specific to the question posed of mutual exclusivity however, there have been a number of papers that have demonstrated that endurance training inhibits the maximum level of strength training that can be developed [4-9]. Indeed it was 20 years ago that a leading article on mutual exclusivity summarised the current understanding at the time that concurrent training had beneficial adaptations in terms of muscle strength and aerobic capacity but that additional endurance work prevented maximal strength acquisition [10]. The postulated mechanisms included both central and peripheral control as well as raising the possibility that it was the training style that may be responsible for this. Although some papers have shown no relevance others more recently have looked at training styles which have been adapted and the phased differently to see if this has any effect supporting the idea of a continuum [11]. Certainly, it is also well shown that circuit training after endurance work does have added benefits [12, 13]. These changes are not specifically age related [14,15]. Also, individuals who partake in any particular sport probably would benefit from the other extreme of their sporting requirements for additional benefits despite the fact that the pathways probably interact [16,17]. Furthermore, de Souza et al. [18] found that concurrent training was not a factor in reducing muscle strength when they compared it to strength training [18]. The ‘motor pathway’ is shown in Figure 1, which demonstrates the gross anatomy involved in causing muscle contraction.
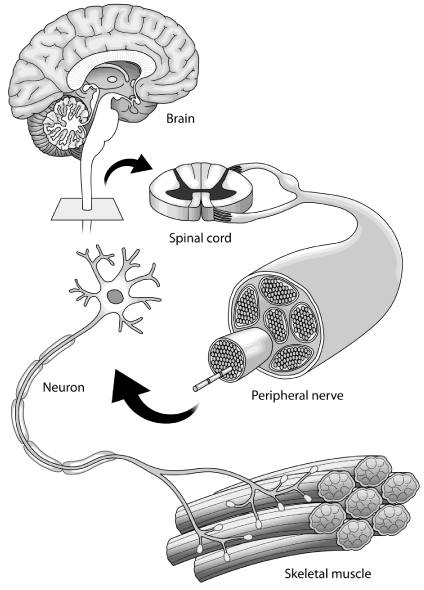
Figure 1: The ‘motor pathway’.
Microscopic
The microscopic changes that occur have shed more light on myofibril adaptation to specific training styles supporting the macroscopic observations above. Again the physiological changes during endurance training are well supported in the literature including general hypertrophy and an alteration from Type II B to Type II A muscle fibres [17,19], increased glycogen and fat storage in the muscle, improved neuromuscular junction and muscle signalling responses improving contraction times and response, mitochondrial hyperplasia and hypertrophy, increased efficiency of mitochondria themselves and increased muscle capillarisation [2,3]. Microscopic strength training changes are however more localised in particular muscle hypertrophy and reduction in aerobic performance [2,3].
The muscle morphological changes occurring through strength and endurance training certainly raise the possibility of these being mutually exclusive, however, research has quite reasonably questioned how this could affect other changes occurring in the endurance group. Certainly changes are specific to the muscles stressed confirming in someway a direct response as part of a muscle specific strength-endurance continuum, rather than mutual exclusivity [20]. Hypotheses raised have also included a central governor control, circulating hormones [21], relevance of insulin control and the question of genotype. The central control is a matter of on-going debate. Neurologically morphological changes occur [22]. Motor units are also summated in different patterns, either spatial (one nerve recruiting more motor units resulting in increased strength) or temporal (increased speed of firing to increase the force of contraction).
Neurologically at the microscopic level it was believed that there were certain learned pathways (engrams) that controlled this. We know from cerebral palsy and developmental conditions that these children lack the ability to memorise these engrams which should be seen a units of electrical code for a particular task. Also there may well be a degree of synaptic plasticity where independent nerve pathways may indirectly affect the result, which was the basis of the Hebbian theory [23].
Research suggests that limiting strength or resistance training can result in exclusive responses; however, linked with macroscopic work this is difficult. As you can imagine, an athlete training for purely endurance work would still walk around, perform activities of daily living (lifting shopping bags, gardening, and sprinting up and down stairs). Any event against gravity will hence impart some degree of strength training without realising. Single-fibre studies have overcome this dilemma, but exclude how muscles are orientated in sheets and angled to their joint line biomechanics, and more central factors of training for example psychology, emotion, stress, noise and ambient temperature [24]. The implication of single-fibre studies is that the muscle response is specific to the nerve stimulating it and therefore muscle responses are exclusive to the nerve stimulus. The fibre then adapts accordingly and specific to the task for its next action with up-stream multilevel memory and protective breaks. Apart from neuromuscular adaptations and the cellular changes, noted above from direct cellular analysis, there is a paucity of other microscopic information comparing the two modes of training. Fat metabolism has also been studied suggesting that endurance athletes handle dietary fat loads differently by moving it more readily into the intramyocellular compartment [25]. There is however a regional and phenotypic variation.
Molecular
The third aspect of this article is on molecular research involving immunohistochemical studies looking at predominantly skeletal muscle plasticity and mitochondrial adaptations, which have unveiled a number of complicated pathways. Evidence for central plasticity is however less well cited regarding exclusivity, however there is one study showing endurance work causing cortical angiogenesis and strength training activating or recruiting spinal motor neurones, and the authors speculate that responses were clearly specific to task [26].
The following papers review muscular adaptations. Spiering et al. [27] produced a theoretical stream of responses and interactions at each level ultimately resulting in muscle adaptation [27]. The paper summarises muscle signalling confirming multiple interactions as well as the role of inhibitory components resulting in a final common step at the last possible point namely the mRNA level. A review article on the molecular basis of training adaptation summarise three postulated pathways for concurrent training responses [28]. These pathways include firstly a calcium-calmodulin dependent kinase pathway acting on a eukaryotic elongation factor 2 kinase (eEF- 2 kinase) resulting in ribosome translation with strength training however inhibiting eEF-2 kinase via S6K. There are a number of types of CaMK and it appears that levels all vary with age and demands [29]. This probably explains the reduction of number of type II fibres with age being genotypically controlled. The alternative pathway is via the FoxO1 transcription factor which after endurance work results in mitochondrial responses but via strength work Akt phosphorylates the FoxO1 factor resulting in protein degradation. Thirdly, Coffey and Hawley mention the most ‘compelling’ mechanism to mediate specificity of training and subsequent interference effect: the AMPK-Akt master switch hypothesis [28]. Endurance work reduces glycogen and increases AMP resulting in AMPK activity and mitochondrial biogenesis. Strength work via the Insulin Growth Factor (IGF) promotes Akt and via the mTOR pathway produces protein synthesis. This particular pathway is important because AMPK also inhibits mTOR, hence interfering with the theoretical response that would be achieved with pure strength training and therefore this is limited [30]. Moreover, Moller et al found that mTORC1 could be activated by IKK? in skeletal muscle, as IKK? phosphorylation is increased with resistance exercise, but not endurance exercise [31]. The molecular basis of strength and endurance training is shown in Figure 2.
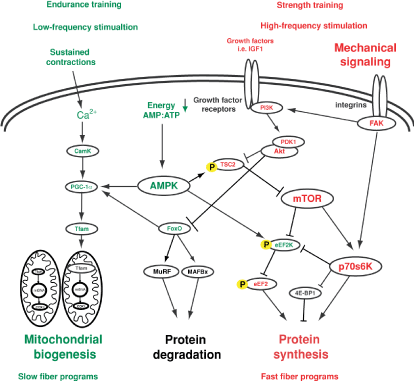
Figure 2: The molecular basis of strength and endurance training. mTOR (strength) and AMPK (endurance) action are the key areas of deviation between strength and endurance training. (Taken, with permission, from: Hoppeler H, Baum O, Lurman G, Mueller M. 2011. Molecular Mechanisms of Muscle Plasticity with Exercise. Comprehensive Physiology. 1383�1412.)
References
- MacAuley D. Oxford handbook of sport and exercise medicine. Oxford, UK: University Press, 2006.
- Brukner P, Khan K. Clinical sports medicine (3rd edn). Sydney, Australia: McGraw-Hill, 2006.
- McArdle W et al. Exercise physiology (6th edn ). Baltimore, USA: Williams and Wilkins, 2007.
- Kraemer WJ, Patton JF, Gordon SE, Harman EA, Deschenes MR, Reynolds K, et al. Compatibility of high-intensity strength and endurance training on hormonal and skeletal muscle adaptations. J Appl Physiol (1985). 1995; 78: 976-989.
- Leveritt M, Abernethy PJ, Barry BK, Logan PA. Concurrent strength and endurance training. A review. Sports Med. 1999; 28: 413-427.
- Rhea MR, Oliverson JR, Marshall G, Peterson MD, Kenn JG, Ayllón FN. Noncompatibility of power and endurance training among college baseball players. J Strength Cond Res. 2008; 22: 230-234.
- Mikkola J, Rusko H, Izquierdo M, Gorostiaga EM, Häkkinen K. Neuromuscular and cardiovascular adaptations during concurrent strength and endurance training in untrained men. Int J Sports Med. 2012; 33: 702-710.
- Wilson JM, Marin PJ, Rhea MR, Wilson SM, Loenneke JP, Anderson JC. Concurrent training: a meta-analysis examining interference of aerobic and resistance exercises. J Strength Cond Res. 2012; 26: 2293-2307.
- Rønnestad BR, Hansen EA, Raastad T. High volume of endurance training impairs adaptations to 12 weeks of strength training in well-trained endurance athletes. Eur J Appl Physiol. 2012; 112: 1457-1466.
- Dudley GA, Fleck SJ. Strength and endurance training. Are they mutually exclusive? Sports Med. 1987; 4: 79-85.
- Rana SR, Chleboun GS, Gilders RM, Hagerman FC, Herman JR, Hikida RS, et al. Comparison of early phase adaptations for traditional strength and endurance, and low velocity resistance training programs in college-aged women. J Strength Cond Res. 2008; 22: 119-127.
- Chtara M, Chamari K, Chaouachi M, Chaouachi A, Koubaa D, Feki Y, et al. Effects of intra-session concurrent endurance and strength training sequence on aerobic performance and capacity. Br J Sports Med. 2005; 39: 555-560.
- Taipale RS, Mikkola J, Nummela A, Vesterinen V, Capostagno B, Walker S, et al. Strength training in endurance runners. Int J Sports Med. 2010; 31: 468-476.
- Verney J, Kadi F, Saafi MA, Piehl-Aulin K, Denis C. Combined lower body endurance and upper body resistance training improves performance and health parameters in healthy active elderly. Eur J Appl Physiol. 2006; 97: 288-297.
- Cadore EL, Izquierdo M, Alberton CL, Pinto RS, Conceição M, Cunha G, et al. Strength prior to endurance intra-session exercise sequence optimizes neuromuscular and cardiovascular gains in elderly men. Exp Gerontol. 2012; 47: 164-169.
- Helgerud J, Rodas G, Kemi OJ, Hoff J. Strength and endurance in elite football players. Int J Sports Med. 2011; 32: 677-682.
- Aagaard P, Andersen JL, Bennekou M, Larsson B, Olesen JL, Crameri R, et al. Effects of resistance training on endurance capacity and muscle fiber composition in young top-level cyclists. Scand J Med Sci Sports. 2011; 21: 298-307.
- de Souza EO, Tricoli V, Roschel H, Brum PC, Bacurau AV, Ferreira JC, et al. Molecular adaptations to concurrent training. Int J Sports Med. 2013; 34: 207-213.
- Jackson CG, Dickinson AL, Ringel SP. Skeletal muscle fiber area alterations in two opposing modes of resistance-exercise training in the same individual. Eur J Appl Physiol Occup Physiol. 1990; 61: 37-41.
- Campos GE, Luecke TJ, Wendeln HK, Toma K, Hagerman FC, Murray TF, et al. Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones. Eur J Appl Physiol. 2002; 88: 50-60.
- Cadore EL, Izquierdo M, dos Santos MG, Martins JB, Rodrigues Lhullier FL, Pinto RS, et al. Hormonal responses to concurrent strength and endurance training with different exercise orders. J Strength Cond Res. 2012; 26: 3281-3288.
- Folland JP, Williams AG. The adaptations to strength training: morphological and neurological contributions to increased strength. Sports Med. 2007; 37: 145-168.
- Paulsen O, Sejnowski TJ. Natural patterns of activity and long-term synaptic plasticity. Curr Opin Neurobiol. 2000; 10: 172-179.
- Malisoux L, Francaux M, Theisen D. What do single-fiber studies tell us about exercise training? Med Sci Sports Exerc. 2007; 39: 1051-1060.
- Tamura Y, Watada H, Igarashi Y, Nomiyama T, Onishi T, Takahashi K, et al. Short-term effects of dietary fat on intramyocellular lipid in sprinters and endurance runners. Metabolism. 2008; 57: 373-379.
- Adkins DL, Boychuk J, Remple MS, Kleim JA. Motor training induces experience-specific patterns of plasticity across motor cortex and spinal cord. J Appl Physiol (1985). 2006; 101: 1776-1782.
- Spiering BA, Kraemer WJ, Anderson JM, Armstrong LE, Nindl BC, Volek JS, et al. Resistance exercise biology: manipulation of resistance exercise programme variables determines the responses of cellular and molecular signalling pathways. Sports Med. 2008; 38: 527-540.
- Coffey VG, Hawley JA. The molecular bases of training adaptation. Sports Med. 2007; 37: 737-763.
- Chin ER. The role of calcium and calcium/calmodulin-dependent kinases in skeletal muscle plasticity and mitochondrial biogenesis. Proc Nutr Soc. 2004; 63: 279-286.
- Nader GA. Concurrent strength and endurance training: from molecules to man. Med Sci Sports Exerc. 2006; 38: 1965-1970.
- Møller AB, Vendelbo MH, Rahbek SK, Clasen BF, Schjerling P, Vissing K, et al. Resistance exercise, but not endurance exercise induces IKKß phosphorylation in human skeletal muscle of training-accustomed individuals. Pflugers Arch. 2013; 465: 1785-1795.