
Research Article
Austin J Plant Biol. 2015; 1(2): 1010.
Restriction of Viscum Album to Few Phorophytes in a Habitat with Diverse Type of Tree Species
Ahmed Z and Dutt HC*
Department of Plant Ecology Laboratory and Botany, University of Jammu, India
*Corresponding author: Dutt HC, Department of Plant Ecology Laboratory and Botany, University of Jammu, Jammu-180006, India
Received: July 27, 2015; Accepted: December 07, 2015; Published: December 16, 2015
Abstract
Viscum known for 464 species is distributed from temperate to tropical world as is a hemi-parasite on tree species. Commonly called as mistletoes, the members of the group show normal parasitic, facultative, or rarely tripartite associations. Mistletoes are also known for auto parasitism, where individuals of the same species parasitize on the same species of mistletoe growing on the host trees. In the present study, V. album is reported on three cultivated host species i.e. Juglans regia L., Prunus armeniaca L. and Salix alba L. only, however, the study area is characterized with the number of other tree species also. In order to understand the specificity of the hemi-parasite on these three phorophytes, a seed germination test of V. album has been conducted on other eight tree species in the same geographical area.
Keywords: Temperate NW himalaya; Parasitism; Phorophyte; Autoparasite; Host specificity
Introduction
Viscum is a hemi-parasite on several tree species and comprises about 464 species widely distributed from temperate to tropical climates in Europe, Africa, Asia, and Australia [1]. Morphologically, V. album is 30–100cm long, dichotomously branched with yellowishgreen, opposite, strap-shaped, entire, leathery leaves measuring 2–8 X 0.8–2.5cm. During reproductive phase species bears insect – pollinated, unisexual, yellowish-green flowers measuring 2–3 mm in diameter. The fruit is white or yellow berries containing one (very rarely several) seed embedded in the very sticky, glutinous fruit pulp [2]. Traditionally, the genus has been placed in family Viscaceae, but on the basis of molecular studies now-a-days all the members of Viscaceae are merged into the Sandalwood family i.e., Santalaceae [3,4]. In general the pattern of host specificity in mistletoes covers a wide spectrum from highly host specific to host generalists [5,6,7]. Some mistletoe parasitizes a large number of hosts in different families. e.g., V. album ssp. album on more than 450 host species and a few mistletoes e.g., some dwarf mistletoes parasitize only one host species [8,9]. The host specificity of dwarf mistletoes may be a useful taxonomic character for distinguishing between host populations [10,11,12,13]. However, some mistletoe is not common on noncultivated plants [14]. In addition, many species have been thought of as key species for animals which eat honey and fruit [15,16]. Another fascinating effect of mistletoe host specificity is the propensity of mistletoes to parasitize mistletoe. This facultative association is termed as hyperparasitism [17]. Hyperparasitic mistletoes are recorded from Loranthaceae and Santalaceae [18]. A number of species of Phoradendron and Viscum also show hyper-parasitism [19,1]. Even more amazing are the rare tripartite associations where a mistletoe parasitizes a mistletoe that is parasitizing another mistletoe on a host tree [18]. When mistletoe on another mistletoe becomes an obligate association, this is termed epiparasitism viz Phacellaria [20,17]. Parasitism by an individual of the same species is called auto parasitism. Auto parasitism occurs frequently in Loranthaceae as well as in Santalaceae (Figure 1).
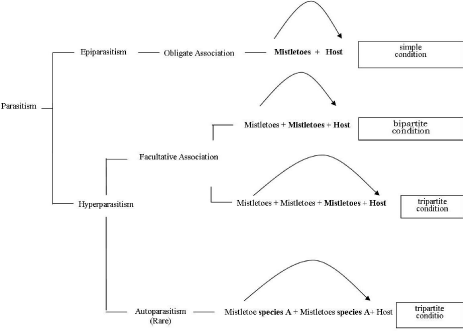
Figure 1: Parasitism.
In the present study we have recorded three phorophytes of Viscum album i.e Juglans regia L, Prunus armanica L and Salix alba Thunb., however, literature review has suggested 452 species as phorophytes, of which Rosaceous has the maximum hosts (128 taxa) infested by V. album [8]. Therefore a study has been conducted to investigate the possibility of new infestations by V. album in the study area.
Material and Methods
Study area
Geographical area flanked between E 75°33/24.92 to N33°08/15.99 and E 75°31/09.41 to N33°01/43.44 and divided in two parts by a perennial stream called as Neeru has been selected for the study. This is included in Bhadarwah Forest Division, Jammu and Kashmir and India is designated as Kellar Forest Range (Figure 2). On the right bank of the stream in this forest range, the mountainous forests are dominating by tree species like Quercus baloot Griff., Punica granatum L., Daphne oleoides Schreb and Olea cuspidata Wall. However, the mountainous slopes in the forest on left bank of the stream comprises mostly conifers like Cedrus deodara Loudon., Pinus roxburgii Sarg., Pinus wallichina A.B. Jacks, Picea smithiana Boiss and Abies pindrow Royle.
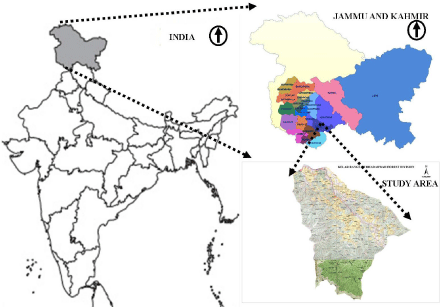
Figure 2: Geographical map showing the study area in northwest Himalaya, Jammu and Kashmir, India.
Test seed germination on different host
To understand the germination of seeds of the V. album on different hosts, seeds of V. album collected in the month of November from natural habitat were implanted on stem of Melia azedarach L., Ficus palmata Forssk., Alnus nitida Endl., Quercus baloot, Diaspyros lotus L., Cedrus deodara, Olea cuspidata and Populus ciliata Wall. The rationality for this experiment was to understand the less distribution and less presence of V. album in the study area and its restriction or specification on Juglans regia L., Prunus armeniaca L. and Salix alba L. only. Percentage of the seed germination has been calculated using formulae
Number of Seeds germinated on tree X 100 Number of Seeds implanted on tree.
Results
The study area is a temperate forest range NW Himalaya between Pir Panjal and Duladhar range. Due to its location on the left bank of river Chenab, it is also called as extension of Pir Panjal (Figure 2). The study area is known for thick coniferous forest and scrub forests on mountainous slopes of left and right bank of Neeru stream, respectively. Among coniferous trees Pinus roxburgii, Cedrus deodara, Pinus wallichiana are found at low elevations and Picea smithiana, Abies pindrow at high elevations. Sporadic distribution of Taxus baccata L., Ficus palmata, Rhododendron arboretum Sm., Viburnum grandiflorum Wall. ex DC., Pyrus pashia Buch.-Ham. ex D. Don and Trema politra (Planch.) Blume is also observed in the study area. Beside the Quercus baloot, Punica granatum, Daphne oleoides and Olea cuspidata other species on the mountains of right bank includes Quercus floribunda Wall and Dodonea viscosa Jacb. However, Alnus nitida Endl. A broad leaved species is very common along riverine ecosystem in the study area. Other common tree species observed in the study area are horticulture (06), timber (02) and fire wood (03) species (Table 1).
![]()
Horticulture species (6)
Timber species (2)
Fire wood species (3)
Pyrus malus
Juglans regia
Salix alba
Prunus armeniaca
Populus cilata
Populus cillata
Juglans regia
Melia azedarach
Pyrus pashia
Cydonia oblonga
Diaspyros lotus
Table 1: Economically important cultivated tree species in the study area.
Despite of the fact that the study area is known for many tree species, V. album has been observed only on three economically important tree species i.e. Juglans regia, Prunus armeniaca and Salix alba. To understand the restricted distribution of the species on few tree species only, field experiment on seed germination of V. album on tree three species has been conducted (Table 2). Seeds of V. album were collected from the field in the month of November- December and implanted on the stem of 08 tree species in January (Table 2). The seeds initiated germination on the stem of 06 species i.e. Melia azedarach, Alnus nitida, Diaspyros lotus, Cedrus deodara, Olea cuspidata and Populus ciliata in the month of February. It has also been observed that implanted seeds of V. album fail to germinate on the stem bark of Ficus palmata and Quercus baloot under same climatic conditions (Table 2 & 3), however they germinate on rest of the species after 50-60 days. Germination percentage calculated for the seeds showed maximum value for Melia azedarach and the lowest percentage for Alnus nitida. The experiment of seed germination has been conducted in the field itself where the air temperature (min/ max) and relative humidity (min/max) has been recorded on daily basis (Table 3). After 120 days of implantation of seeds on 08 different species, no penetration of the haustoria of germinated seeds V. album has been recorded on any of the tree species.
![]()
Name of the phorophyte/ plant model
Usage and type of resource
Life form of Phorophyte/ plant model
Bark texture of Phorophyte/ plant model
% age of seed germination of V. album on plant model
Status of the implanted seeds of V. album on plant model after 60 days
Status of the implanted seeds of V. album on plant model after 120 days
Juglans regia *
H,c
Tree
Furrowed
NA
NA
NA
Prunus armeniaca *
H,c
Tree
Fissured
NA
NA
NA
Salix alba*
FW,c
Tree
Fissured
NA
NA
NA
Ficus palmata†
F,n
Tree
Fissured
0
_
Olea cuspidata †
F,n
Tree
Fissured
50
A
nphb
Populus ciliata †
T,c
Tree
furrowed
20
A
nphb
Quercus baloot †
FW,n
Tree
Scaly
0
_
Alnus nitida†
FW,n
Tree
Smooth
16.6
A
nphb
Diaspyros lotus†
H,c
Tree
Smooth
30
A
nphb
Cedrus deodara †
T,n
Tree
Smooth
40
A
nphb
Melia �azedarach †
FW,c
Tree
Smooth
60
A
nphb
* = Natural phorophyte; † = plant model; H= horticulture tree; FW = Fuel wood ; F = Fodder; T= Timber wood; c= cultivated tree; n= natural tree; A=alive, nphb = not penetrated in host bark
Table 2: Natural phorophytes and testing of host specificity of Viscum album.
![]()
Months
Air Temperature
Relative Humidity
Minimum
Maximum
Minimum
Maximum
February
6.2
15.6
46.1
70.4
March
8.8
19.1
43.5
67.0
April
14.9
23.5
43.9
66.5
May
16.3
25.6
45.2
60.3
June
18.0
30.4
53.6
61.5
Table 3: Mean value of the air temperature and relative humidity during study period.
Discussion
V. album is a partial parasite on tree species and distributed almost all over the world. About 452 tree species are known as the successful host of the species and Rosaceous is considered as the most frequent host with large number of the tree species as phorophytes. However, commonly infested genera include Salix, Populus, Acer, Malus, Prunus, Sorbus, Abies and Pinus [21]. It is understood that seed of many plant parasites germinates only in response to chemical signals from host plants [21]. In our study V. album is observed on three cultivated hosts i.e. Juglans regia, Prunus armeniaca or Salix alba, this indicates that the parasite is introduced to the study area along with the cultivated tree species. However, simultaneously no temporal data on the introduction of the host species is available. To understand the host specificity and less distribution of V. album on other tree species, seed germination test for V. album has been conducted on 03 cultivated and 05 naturally occurring tree species in the study area (Table 1). These tree species are cultivated for number of reasons like Populus ciliata for timber, Diaspyros lotus as horticultural crop and Melia azedarach as fuel wood tree. Variability in the bark texture has been maintained among the tree species subjected for the experimentation, 04 species (Alnus nitida, Diaspyros lotus, Cedrus deodara and Melia azedarach) were having smooth bark, 01 each was having scaly (Quercus baloot) and furrowed (Populus ciliata) and 02 were having fissured bark (Olea cuspidata, Ficus palmata). During the experiment conducted on seed germination on bark of different tree species maximum seed germination percentage (60%) has been observed on Melia azedarach followed by Olea cuspidata (50%), Cedrus deodara (40%), Diaspyros lotus (30%), Populus ciliata (20%) and Alnus nitida (16.6 %), however seeds were not germinated on Ficus palmata and Quercus baloot (Table 2). This indicates that type of the phorophyte provide stimulus to germinate the seeds of parasite on it, this is in consonance with the statement made on germination of seeds of the parasites on host only in response to chemical signals from host plants [21]. However, some mistletoe seeds germinate readily in almost all situations. The key limiting step in mistletoe’s life cycle is establishment, which is dependent on an appropriate disperser, deposition on a suitable sized branch, and mistletoe-host compatibility [22,23]. The random germination on each type of bark suggests that there is no role of the type of bark on the germination of the seeds. The role of bark texture may be providing a suitable site for seeds when they get shed from the parent plant. The rough barks are more appropriate as they provide the best platform in the fissures and grooves for the settlement of the seeds, e.g., Juglans, Salix and Prunus.

Figure 3: Geographical map showing the study area in northwest Himalaya, Jammu and Kashmir, India.
The exact stimulus for the germination of the seeds on a particular phorophyte is not clear, i.e., either controlled by the phorophyte itself or by environmental factors. In our experiment, as stated in the results the seeds of V. album germinates on six phorophytes viz. Melia azedarach, Alnus nitida, Diaspyros lotus, Cedrus deodara, Olea cuspidata and Populus ciliata and was not able to germinate and the rest two hosts i.e., Quercus baloot and Ficus palmata, indicates that the seeds of V. album do not germinate on all the tree species under same climatic condition and same geographical area (Table 3). Ethno-botanically the species is used to treat the hypertension [24]. In accordance with Musselman and Press, 1995, seed germination of parasitic plants is dependent upon the chemical signals from the host however we suggest that for establishment of the parasitic species, chemical signal studies are required to be conducted. Because, the seed germination in initial 60 days is observed as normal process however, establishment of the parasite even after 120 days of implantation has not be found in the study area under same climatic condition, where the Viscum album is reported infesting three tree species naturally.
Conclusion
Beside chemical signal from host species for germination of seeds of V. album, a chemical signal for establishment of the germinated seeds may are also be required from host plant, so that the parasite can establish on its host. Otherwise the germinated seeds cannot get established on host.
References
- Polhill R, Wiens D. Mistletoes of Africa. The Royal Botanic Gardens, Kew. 1998; 370.
- Zuber D. Biological flora of Central Europe: Viscum album L. Flora. 2004; 199: 181-203.
- Anonymous. An update of the Angiosperm Phylogeny group classification for the orders and families of flowering plants: APG II. Bot J Linn Soc. 2003; 141: 399-436.
- Chase MW, Reveal JL. A Phylogenetics classification of the land plants to accompany APG-III. Bot J Lin Soc of London. 2009; 161: 122-127.
- Barlow BA, Wiens D. Host-parasite resemblance in australian mistletoes: the case for cryptic mimicry. Evolution. 1997; 31: 69-84.
- Mellado A, Zamora R. Linking safe sites for recruitment with host-canopy heterogeneity: The case of a parasitic plant, Viscum album subsp. austriacum (Viscaceae). Am J of Bot. 2014; 101: 957-964.
- Zuber D. Widmer A. Genetic evidence for host specificity in the hemi-parasitic Viscum album L. (Viscaceae). Molecular Ecology. 2000; 9: 1069-1073.
- Barney CW, Hawksworth FG, Geils BW. Hosts of Viscum album. Eur J For Path. 1998; 28: 187-208.
- Geils BW, Hawksworth FG, Damage, effect and importance of dwarf mistletoes, In: Mistletoes of North America Conifers. USDA. 2002; 57-65.
- Aukema JE. Vectors, viscin and Viscaceae: mistletoes as parasites, mutualists, and resources. Frontiers in Ecology and the Environments. 2003; 1: 212-219.
- Hawksworth FG, Scharpf RF, Marosy M. European mistletoe continues to spread in Sonoma County. California- Agriculture. 1991; 45: 39-40.
- Hawkswort, FG. Coevolution of Mexican white pines and their dwarf mistletoe parasites. In: Proceedings, symposium on white pine provenances and breeding Montreal, Quebec. Gen. Tech. Rep. NE-155. Radnor, PA: U.S. Department of Agriculture, Forest Service, Northeast Forest Experiment Station. 1991; 23–28.
- Mathiasen RL, Daugherty CM. Distribution of red fir and noble fir in Oregon based dwarf mistletoe host specificity. Northwest Sci. 2008; 82: 108-119.
- Silva A, Rio MD. Effects of the mistletoe Tristerix aphyllus (Loranthaceae) on the reproduction of its cactus host Echinopsis chilensis. Oikos. 1996; 75: 437–442.
- Morand S, Krasnov BR. The Biogeography of host-parasite interactions, Oxford University Press, New-York. 2010; 288.
- Watson DM. Mistletoe- A keystone resource in forests and woodland and worldwide. Annual Review of Ecology and Systematic. 2001; 32: 219-249.
- Visser JH. Epiparasitism – how many mistletoes on one another? Golden Baugh (Royal Botanic Gardens, Kew). 1982; 1: 4.
- Nickrent DL. Parasitic plants of the world. in: plantus parasites del la peninsula iberica e islas (Eds. Baleares JA, Lopez Saez PC, Rodriguez, Saezeds L.) Mundipresna, Madrid, Spain. 2002; 7-24.
- Kuijt J. Monograph of Phoradendron (Viscaeace). Syst Bot Monogr. 2003; 66: 1-643.
- Muir JA, Geils BW. Management strategies for dwarf mistletoes: Silviculture. In: Proceedings, symposium on white pine provenances and breeding Montreal, Quebec. Gen Tech Rep. NE-155. Radnor, PA: U.S. Department of Agriculture, Forest Service, Northeast Forest Experiment Station. 1991; 83-94.
- Musselman LJ, Press MC. Introduction to parasitic plants. In: Press MC, Graves JD, eds. Parasitic plants. London: Chapman and Hall. 1995; 1-13.
- Norton DA, Reid N. Lessons in ecosystem management from management of threatened and pest Loranthaceous mistletoes in New Zealand and Australia. Conservation Biology. 1997; 11: 759-769.
- Reid N, Smith NM, Yan Z. Ecology and population biology of mistletoes. – In: Forest canopies. Academic Press (Eds. Lowman MD, Nadkarni NM), 1995: 285–310.
- Dutt HC, Bhagat N, Pandita S. Oral traditional knowledge on medicinal plants in jeopardy among Gaddi shepherds in hills of northwestern Himalaya, J&K and India. Journal of Ethno pharmacology. 2015; 168: 37–348.