
Mini Review
Thromb Haemost Res. 2017; 1(2): 1008.
Mechanisms of Platelet Activation, Adhesion and Aggregation
Hosseinzadegan H and Tafti DK*
Mechanical Engineering Department, Virginia Polytechnic Institute and State University, USA
*Corresponding author: Tafti DK, Mechanical Engineering Department, Virginia Polytechnic Institute and State University, 635 Prices Fork Rd, Blacksburg, VA 24060, USA
Received: November 21, 2017; Accepted: December 14, 2017; Published: December 28, 2017
Abstract
In this article, observed mechanisms of platelet activation, adhesion and aggregation at the molecular level are reviewed. The role of different blood constituents, platelet integrins, tissue factors, and extracellular matrix ligands in platelet adhesion and activation are also presented. In extreme flows, as found in heart assist devices or severely stenosed vessels, pathologic platelet deposition is very likely to occur. For this reason, experimental measurements on effects of flow characteristics, particularly shear stress, on the rate of platelet accumulation, embolism, and thrombus stability are also reviewed in this paper.
Keywords: Glycoprotein; Hemostasis; Integrin; Shear stress; Thrombosis, von Willebrand factor
Abbreviations
ADP: Adenosine Diphosphate; CICR: Calcium Induced Calcium Release; ECM: Extracellular Matrix; GP: Glycoprotein; GPCR: G Protein-Coupled Receptor; PPACK: D-Phe-Pro-Arg Chloromethylketone; TP: Thromboxane receptor on Platelet; TxA2: Thromboxane A2; VLA: Very Late Antigen; vWD: von Willebrand Disease; vWf: von Willebrand factor
Introduction
Platelets whether suspended in the blood stream or attached to the vessel wall are the main blood component involved in blood clot formation [1-3]. They can bind together and have three forms depending on the flow conditions: grouping of platelets in hemostasis called plug, floating platelet thrombi (aka emboli) that can be carried by blood flow, and deposited mural platelet aggregates on sub endothelium known as plaque. Hemostasis is a natural mechanism, which arrests hemorrhage [4]; however the other two forms occur only under pathologic conditions. Several studies in various vasculatures have been carried out to elucidate the role of potential mechanical and biological elements at play. The role of platelet activation in adhesion, platelet adhesion rate, and the function of different molecules and cells are dictated by the biophysics of the blood flow. Therefore, processes of flow and mass transfer strongly affect the platelet accumulation rate, intraclot transport, and clot stability and decomposition.
Platelet activation and its role in adhesion and aggregation
Once platelets adhere to the vessel wall, specifically to the endothelium, they get activated, change dramatically in shape from a discoid ball to a sphere [5] to first increase the adhesion area and second, stimulate other platelets resting in plasma to become activated through biological mechanisms. These mechanisms include signaling pathways [6] in the platelet extracellular membrane and secretion of soluble thrombogenic agonists into the plasma. Activated platelets in turn release their stored granules, including ? granules and dense granules to the plasma, which leads to sudden increase in the local concentration of agonists necessary for atherosclerosis [7]. Unlike the initiation of adhesion of platelets exposed to subendothelium, the formation of platelet aggregates is based on the platelet-platelet and platelet-agonists interactions.
Constituents involved in platelet activation
Collagen, Adenosine Diphosphate (ADP), Thromboxane A2 (TxA2), thrombin, and fluid shear stress can alter processes leading to platelet activation. Therefore, platelet activation cannot be determined solely by one of these factors. However, each factor has its own contribution and could be defining for activation. Collagen can activate platelets upon surface contact [8] and the activation is mediated by Gly-Pro-Hyp sequence and requires the presence of glycoprotein VI on the platelet membrane [9].
The role of ADP in platelet activation has been studied broadly to identify its contribution to the process [10-14]. Platelet activation propagation away from the wall is attributed to ADP proteins, which are released by dense granules from activated platelets [15]. So, ADP makes activation possible for platelets without their direct contact with the vessel wall, but it has been shown that it has a major role only at low concentrations (<1μM)[16]. Thrombin (aka coagulation factor IIa or E.C.3.4.21.5) can form through proteolysis process and transformation of factor II (prothrombin) to IIa during platelet activation. The role of thrombin in activation has been quantified [17,18] and compared to collagen-induced activation by [19]. Enough concentration of collagen is required for thrombin generation from prothrombin (referred to as the threshold effect of collagen). In the presence of 4 μ/ml collagen, 0.1 nmol/L thrombin is enough to evoke maximal platelet activity in prothrombin and factor X activation [20]. In the absence of collagen, there is a slow increase in both activities for increased amount of thrombin. For high thrombin concentrations, the platelet activity is still lower that of platelets stimulated by the combined action of thrombin and collagen. It was shown that the collagen concentration of 0.5, 0.75 nmol/L and 1.2 nmol/L, respectively have threshold, half-maximal, and maximal effects in platelet activation and aggregation. For collagen concentrations higher than 1.2 nmol/L, the generation of thrombin is not affected by collagen (asymptotic effect of collagen). Platelet activation also stimulates formation of another aggregation agent that contributes to thrombus formation, TxA2 with platelet signaling via the arachidonic pathway [18,21,22]. TxA2 along with thrombin and ADPs bind to their specific receptors on the platelet membrane and activate those receptors [23]. The formation of microparticles is another implication of platelet activation and is widely used as a detector of platelet activation in experiments [24,25]. Figure 1 summarizes the major components involved in platelet activation and adhesion [26].
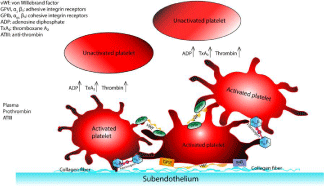
Figure 1: Blood constituents and tissue factors involved in platelet activation
and adhesion. Images are reprinted from [26] and with permission from Wiley
Online Library.
Platelet adhesion to subendothelium
The role of integrins and receptors: In case of occurrence of pathological changes, platelets tend to adhere to the subendothelium to arrest the bleeding and maintain the vessel wall integrity. One of the prerequisites is the presence of specific receptors on platelets and vessel wall. Both vessel wall and platelets’ surface have integrins and receptors on them.
Subendothelium-specific integrins: Collagen is one of the major thrombogenic proteins on the vessel walls, available in the subendothelial matrix [27]. The integrin a2β1 and the glycoprotein GPVI are the adhesion receptors of collagen [28,29]. GPVI is the common adhesion receptor between collagen and platelets. The von Willebrand (vWf) protein, synthesized in the platelet’s internal cytoskeleton core (megakaryocytes) [30] and in the endothelial cells [31], is the other factor available in soluble state both on vessel walls and in plasma. The role of vWf in mediating platelet adhesion was first indicated by [32]; 10 times longer bleeding time caused by impaired platelet adhesion to vessel wall surface was shown for patients with von Willebrand disease (vWD). The relationship between the levels of human vWf and platelet adhesion to cultured endothelial cells under ex-vivo and in-vivo conditions has been established in pig lung, porcine pulmonary artery and aorta [33,34]. In addition to these components, fibronectin, thrombospondin, and laminin from the subendothelium could affect the rate of platelet adhesion to the subendothelial matrix [5,35]. The role of Glycoprotein IV (GPIV) is unclear as the other collagen receptor [8,15]; it can probably mediate adhesion [36]. However, Tandon et al. [37] showed that this glycoprotein can adversely affect platelet activation as 10 nM of GPIV resulted in 50% inhibition of collagen-induced platelet activation.
Platelet-specific receptors
The integrins a2β1 (referred to as GPIa-IIa), Talin-1, GPVI, GPIb-IX-V and vWf are the five adhesion receptors available on the platelet membrane surface [30,38]. Talin-1 is the essential integrin in the platelet adhesion process [23]. Also GPIa has been shown to be necessary for human platelet-collagen adhesion [34]. Platelets can have the vWf on their outer surface [39], being formed by glycoprotein GPlb-IX-V [30]. The other adhesive integrin receptor, aIIbβ3 (known as GPIIb-IIIa or fibrinogen receptor) available on the platelet extracellular membrane [28], enables platelet-platelet binding and comes into play when an activated platelet is exposed to adjacent activated platelets and fibrinogen proteins floating in the plasma [11]. The role of aIIbβ3 in initiation and early formation of platelet adhesion to the subendothelial cells under in-vivo conditions is not clear yet; however, studies suggest that GPIIb-IIIa along with a5β1 (VLA-5) is one of the two necessary factors in mediating platelet adhesion to a fibronectin-coated surface [15,40].
Platelets mainly adhere to collagen (types I, III, IV) or to vWf molecules found in collagen [15,41]. Collagen fibers can augment adhesion by connecting to platelet glycoprotein, GPIa-IIa. GPIa-IIa is the most important receptor for collagen also known as a2β1 or VLA- 2 [40] and binds to the Extracellular Matrix (ECM) components [28] and mediates adhesion of all types of collagen [40-42]. Platelets also adhere to collagen via GPVI and to vWf via the membrane adhesion receptor, glycoprotein Ib-IX-V (GPlb-X-V) [28]. vWf in turn can bind to fibronectin-containing fibers [41], collagen types IV and V and purified collagen types I and III [31]. Studies suggest that the specific component of the subendothelium to which vWf binds is type VI collagen [43]. Platelets can also adhere to type I collagen fibrils with bound vWf on the surface tissue with mediation of platelet integrins a2β1 and aIIbβ3 [44]. In addition to vWf, laminin, fibronectin, and thrombospondin on the vessel surface may play a role, although minor [35], in mediating platelet adhesion [15,23,40,45]. Table 1 lists different matrix and blood components and their associated platelet receptors.
![]()
Soluble agonists
Platelet receptors
ADP
P2X1, P2Y1, P2Y12
Thrombin
GPIba, PAR1, PAR4
TxA2
TP
Epinephrine
a2A
Subendothelial matrix ligands
Platelet receptors
vWf
GPIb-V-IX
Collagen
a2Β1
Fibronectin
a5Β1
Collagen, Laminin
GPVI
vWf, fibrinogen, fibronectin
aIIbΒ3
Table 1: Blood and subendothelial matrix constituents and their associated platelet receptors. In the extracellular space, the soluble agonists bind their cognate platelet receptors to initiate inside-out signaling. Binding of platelet adhesion receptors to subendothelial matrix ligands (collagen, vWf, laminin, fibronectin) or to immobilized fibrinogen deposited from the lumen area initiate the signaling pathways.
Floppy vs. firm adhesion
Platelet adhesion to the Extracellular Matrix (ECM) can be floppy (at initial stage) or firm depending on the type of participating receptors (Figure 2). For reversible platelet adhesion or tethering, the interaction between vWf from exposed subendothelium collagen and the platelet receptor GPIab (GPIb-V-IX) [23] through signaling pathways [6] or presence of enough platelet vWf integrin [39] is required. This leads to unstable adhesion [44], whereas firm platelet adhesion on Extracellular Cell Matrix (ECM) or collagen is mediated by the platelet receptor GPVI together with other platelet receptor G Protein-Coupled (GPCRs) stimulated by local secretion of agonists [23]. Along with these stimulated receptors, the platelet integrin β1 converts from low affinity resting state to high affinity state and binds to fibronectin, laminin collagen, and aIIbβ3. Among these receptors, aIIbβ3 is the key element on platelet membrane required for firm adhesion. Calcium Induced Calcium Release (CICR) mechanism also sustains and amplifies integrin aIIbβ3 activation resulting in firm adhesion [6].
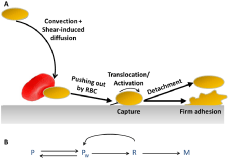
Figure 2: (A) Stages of platelet adhesion and activation [46]. A traveling
platelet is pushed toward the wall by shear-induced diffusion mechanism
and captured at the wall. Irreversible platelet adhesion occurs if platelet is
decelerated and activated; otherwise flow will detach the captured platelet.
(B) The associated kinetic scheme; P: free platelet transported by the flow
far from the wall; Pw: the same platelet in the near-wall layer; R: the captured
platelet rolling along the wall; M: firmly adhered platelet.
Effect of blood flow and shear rate
The contribution of interactions between specific adhesive receptors and soluble stimuli to thrombogenesis can be dependent on the blood flow conditions [46]. Although the sequence of events associated with platelet adhesion, aggregation and activation may remain the same, their occurrence and intensity, as well as the interaction of contributing components with each other can be highly dependent on the local blood flow characteristics [47].
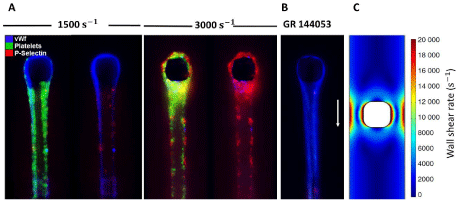
Figure 3: vWf, platelets, and P-Selectin visualized in a small post located in the middle of flow within a microfluidic channel. Platelets are activated and adhere to
fibrous vWf which was generated from initial fiber capture using apixaban and PPACK-inhibited platelet-free plasma at shear rate of 10,000 sec-1. Then, vWf was
labeled using polyclonal fluorescent anti-vWf antibody in HBS and then was washed with HBS to reduce background. The right and left images at each shear rate
are shown to highlight the activated platelets on the surface of vWf. (A) The shear rate was reduced to 1500 sec-1 and 3000 sec-1 and apixaban and PPACK-inhibited
whole blood labeled with platelets and P-selectin was perfused on fibrous vWf. Platelets attach, roll and firmly bind to the fibrous vWf at both shear rates. (B) The
addition of GR 144053 (aIIbβ3 antagonist) to whole blood leads to inhibition of firm platelet adhesion to the vWf fibers. (C) Wall shear rate contours around the post
at the flow rate of 5 μL/min. Images are reprinted from [55] and with permission from Wiley Online Library.
Platelet adhesion and aggregation take place under a wide range flow conditions (shear rates of 20-1,800 sec-1) as well as at shear rates as high as 20,000 sec-1 [48]. The effect of flow shear rate on rate of platelet adhesion to subendothelium was first investigated extensively in the literature by [49-51]. The initial rate of platelet adhesion was shown to increase with increasing vessel wall shear rate up to shear conditions as high as 2,600 sec-1. After 2,600 sec-1, the rate of adhesion decreased with increasing wall shear rate. It has also been indicated that thrombi formation rate increases suddenly between shear rates of 650 sec-1 and 1,300 sec-1 and continues increasing up to a shear rate of 10,000 sec-1 [52].
Several studies have been performed to reveal the potential relationship between local shear stress in the blood flow and unfavorable platelet aggregation and adhesion in various vascular systems. Bark et al. [53] experimentally studied the platelet adhesion in stenosed tubes and reported an increase in platelet adhesion with increasing wall shear rate from 100 sec-1 to 6000 sec-1. After 6000 sec-1, they observed a negative correlation between platelet adhesion rate and wall shear rate. Mehrabadi et al. [54] reported a similar trend of platelet adhesion; however they reported maximum platelet adhesion at a shear rate of 20,000 sec-1. They also attributed the rapid thrombus growth at high shear rates (>4000 sec-1) to elongation of vWf which mediates platelet capture. The contribution of interactions between specific adhesive receptors and soluble stimuli to thrombogenesis can be dependent on the blood flow conditions. Thus conclusions based on experiments carried out under high shear conditions may not necessarily generalized to low shear conditions [6].
Platelet activation also requires a certain critical level of shear rate to occur. In arterial flows, platelet activation is reported for shear rate values of higher than 400~500 sec-1 [24]. This activation increases significantly in higher shear rates (>10,500 sec-1).
Interactions between adhesive receptors on arterial extracellular matrix and platelets, and their subsequent activation and response, required for platelet adhesion, are also functions of blood flow conditions. Therefore, understanding the shear-dependent phenomena associated with the response of these factors in different stress conditions is necessary to explain the dependence of reaction kinetics as well as platelet adhesion, activation, and aggregation on shear stress. The strong dependence of vWf on shear rate, as one of the most important adhesive glycoproteins, was explained by [15]; in flow conditions with shear rates lower than a critical threshold, adhesion of platelets takes place by mechanical pathways, such as diffusion, rather than chemical reaction kinetics [15,52]. However, in high shear rates platelet adhesion to subendothelium is mediated by surface reactivity of platelets, which requires necessary amounts of vWf molecules. Besides, the intensity of platelet adhesion to vWf-rich subendothelium is directly correlated with local shear rate. Herbig and Diamond [55] showed that platelets roll and adhere on vWf fibers at both shear rates of 1500 sec-1 and 3000 sec-1, but pathological shear rate (3000 sec-1) results in the generation of P-Selectin (Figure 3). P-selectin functions as a cell adhesion molecule on the surfaces of activated endothelial cells. This matches with previous observations on endothelial-anchored vWf [56], immobilized plasma vWf [57,58], and fibrous vWf on collagen [59] and shows the important role of fibrous vWf in shear-mediated platelet activation.
Activation of integrin, aIIbβ3, the most important integrin that mediates platelet firm attachment, also depends on the acting shear rate. At .γ < 200 sec-1, a2Β1 and aIIbβ3 get activated by bonds between GPVI receptors and collagen so fast that platelets firmly stick to the surface of the vessel wall.
The shear-dependent role of thrombin was also studied by [60]. It has been indicated that thrombin contributes to thrombus formation only in low shear rate conditions (<650 sec-1) or shear rates higher than 2600 sec-1 with perfusion times as long as 30 min.
Activation-independent platelet adhesion
Although binding of adjacent activated platelets lead to formation of aggregates, platelet aggregation, as well as stable adhesion, does not necessarily require their activation [57]. Under high shear conditions platelets can aggregate without requiring platelet activation. Platelet aggregates can form in the lumen area without needing to have contact with the vascular surface when resting platelets are subject to very high fluid shear rates (>5,000 sec-1) [61].
Conclusion
Blood components, including platelets, and surface tissue factors operate under a wide range of flow conditions, from stasis to depressed flows, from venous to arterial flows, to extreme flows of stenosis or other pathologies. Molecular and cellular mechanics, as well as biochemical reaction kinetics, have been studied in the context of the macroscopic flow hemodynamics. Numerous studies exist for modeling and in vitro and in vivo investigation of blood components function under relevant flow conditions [48,51,62-65]. Further research in this area will help identify patients with high risk of thrombosis and designing improved diagnosis and assist devices [66,67].
References
- De Gaetano G. Historical overview of the role of platelets in hemostasis and thrombosis. Haematologica. 2001; 86: 349–356.
- Monroe DM, Hoffman M, Roberts HR. Platelets and thrombin generation. Arterioscler Thromb Vasc Biol. 2002; 22: 1381–1389.
- Ruggeri ZM. Platelets in atherothrombosis. Nat Med. 2002; 8: 1227–1234.
- Clemetson KJ. Platelets and primary haemostasis. Thromb Res. 2012; 129: 220–224.
- Vorchheimer DA, Becker R. Platelets in atherothrombosis. Mayo Clinic Proceedings. Elsevier. 2006; 81: 59–68.
- Jackson SP, Nesbitt WS, Kulkarni S. Signaling events underlying thrombus formation. J Thromb Haemost. 2003; 1: 1602–1612.
- Huo Y, Ley KF. Role of platelets in the development of atherosclerosis. Trends Cardiovasc Med. 2004; 14: 18–22.
- Siljander P, Lassila R. Studies of Adhesion-Dependent Platelet Activation Distinct Roles for Different Participating Receptors Can Be Dissociated by Proteolysis of Collagen. Arterioscler Thromb Vasc Biol. 1999; 19: 3033–3043.
- Knight CG, Morton LF, Onley DJ, Peachey AR, Ichinohe T, Okuma M, et al. Collagen–platelet interaction: Gly-Pro-Hyp is uniquely specific for platelet Gp VI and mediates platelet activation by collagen. Cardiovasc Res. 1999; 41: 450-457.
- Woulfe D, Yang J, Brass L. ADP and platelets: the end of the beginning. J Clin Invest. 2001; 107: 1503-1505.
- Frojmovic M, Wong T, van de Ven T. Dynamic measurements of the platelet membrane glycoprotein IIb-IIIa receptor for fibrinogen by flow cytometry. I. Methodology, theory and results for two distinct activators. Biophys J. 1991; 59: 815-827.
- Moake JL, Turner NA, Stathopoulos NA, Nolasco L, Hellums JD. Shear-induced platelet aggregation can be mediated by vWF released from platelets, as well as by exogenous large or unusually large vWF multimers, requires adenosine diphosphate, and is resistant to aspirin. Blood. 1988; 71: 1366–1374.
- Hubbell JA, McIntire L V. Visualization and analysis of mural thrombogenesis on collagen, polyurethane and nylon. Biomaterials. 1986; 7: 354–363.
- Puri RN, Colman RW, Liberman MA. ADP-lnduced Platelet Activation. Crit Rev Biochem Mol Biol. 1997; 32: 437–502.
- Weiss HJ. Flow-related platelet deposition on subendothelium. Thromb Haemost. 1995; 74: 117–122.
- Jung SM, Moroi M. Signal-transducing mechanisms involved in activation of the platelet collagen receptor integrin α2β1. J Biol Chem. 2000; 275: 8016–826.
- Fitzgerald DJ, FitzGerald GA. Role of thrombin and thromboxane A2 in reocclusion following coronary thrombolysis with tissue-type plasminogen activator. Proc Natl Acad Sci. 1989; 86: 7585–7589.
- Wagner WR, Hubbell JA. Local thrombin synthesis and fibrin formation in an in vitro thrombosis model result in platelet recruitment and thrombus stabilization on collagen in heparinized blood. J Lab Clin Med. 1990; 116: 636–650.
- Nieuwenhuis HK, Akkerman JWN, Houdijk WPM, Sixma JJ. Human blood platelets showing no response to collagen fail to express surface glycoprotein Ia. Nature. 1985; 318: 470–472.
- Rosing J, Van Rijn JL, Bevers EM, Van Dieijen G, Comfurius P, Zwaal RF. The role of activated human platelets in prothrombin and factor X activation. Blood. 1985; 65: 319-332.
- Clemetson KJ, Clemetson JM. Platelet receptor signalling. Hematol J Off J Eur Haematol Assoc. 2004; 5: 159-163.
- Golino P, Buja LM, Ashton JH, Kulkarni P, Taylor A, Willerson JT. Effect of thromboxane and serotonin receptor antagonists on intracoronary platelet deposition in dogs with experimentally stenosed coronary arteries. Circulation. 1988; 78: 701–711.
- Varga-Szabo D, Pleines I, Nieswandt B. Cell adhesion mechanisms in platelets. Arterioscler Thromb Vasc Biol. 2008; 28: 403–412.
- Sakariassen KS, Holme PA, Orvim U, Barstad RM, Solum NO, Brosstad FR. Shear-induced platelet activation and platelet microparticle formation in native human blood. Thromb Res. 1998; 92: 33–41.
- Holme PA, Orvim U, Hamers MJAG, Solum NO, Brosstad FR, Barstad RM, et al. Shear-induced platelet activation and platelet microparticle formation at blood flow conditions as in arteries with a severe stenosis. Arterioscler Thromb Vasc Biol. 1997; 17: 646–653.
- Hosseinzadegan H, Tafti DK. Modeling thrombus formation and growth. Biotechnol Bioeng. 2017; 114: 2154-2172.
- Coller BS, Beer JH, Scudder LE, Steinberg MH. Collagen-platelet interactions: evidence for a direct interaction of collagen with platelet GPIa/IIa and an indirect interaction with platelet GPIIb/IIIa mediated by adhesive proteins. Blood. 1989; 74: 182–192.
- Gawaz M, Langer H, May AE. Platelets in inflammation and atherogenesis. J Clin Invest. 2005; 115: 3378-3384.
- Nieswandt B, Watson SP. Platelet-collagen interaction: is GPVI the central receptor? Blood. 2003; 102: 449–461.
- Patel SR, Hartwig JH, Italiano Jr JE. The biogenesis of platelets from megakaryocyte proplatelets. J Clin Invest. 2005; 115: 3348-3354.
- Weiss HJ. von Willebrand Factor and Platelet Functiona. Ann N Y Acad Sci. 1991; 614: 125–137.
- Tschopp TB, Weiss HJ, Baumgartner HR. Decreased adhesion of platelets to subendothelium in von Willebrand’s disease. J Lab Clin Med. 1974; 83: 296–300.
- Bahnak BR, Wu Q, Coulombel L, Assouline Z, Kerbiriou-Nabias D, Piétu G, et al. Expression of von Willebrand factor in porcine vessels: heterogeneity at the level of von Willebrand factor mRNA. J Cell Physiol. 1989; 138: 305–310.
- Tersteeg C, De Meyer SF, Deckmyn H. Inhibitors of platelet adhesion to VWF and collagen. Platelets in Thrombotic and Non-Thrombotic Disorders. Springer. 2017; 1313–1323.
- Houdijk WP, De Groot PG, Nievelstein PF, Sakariassen KS, Sixma JJ. Subendothelial proteins and platelet adhesion. von Willebrand factor and fibronectin, not thrombospondin, are involved in platelet adhesion to extracellular matrix of human vascular endothelial cells. Arterioscler Thromb Vasc Biol. 1986; 6: 24–33.
- Tandon NN, Ockenhouse CF, Greco NJ, Jamieson GA. Adhesive functions of platelets lacking glycoprotein IV (CD36). Blood. 1991; 78: 2809–2813.
- Tandon NN, Kralisz U, Jamieson GA. Identification of glycoprotein IV (CD36) as a primary receptor for platelet-collagen adhesion. J Biol Chem. 1989; 264: 7576–7583.
- Clemetson KJ, Clemetson JM. Platelet collagen receptors. Thromb Haemost. 2001; 86: 189–197.
- Kulkarni S, Dopheide SM, Yap CL, Ravanat C, Freund M, Mangin P, et al. A revised model of platelet aggregation. J Clin Invest. 2000; 105: 783-791.
- Beumer S, IJsseldijk MJ, de Groot PG, Sixma JJ. Platelet adhesion to fibronectin in flow: dependence on surface concentration and shear rate, role of platelet membrane glycoproteins GP IIb/IIIa and VLA-5, and inhibition by heparin. Blood. 1994; 84: 3724–3733.
- Wagner DD, Urban-Pickering M, Marder VJ. Von Willebrand protein binds to extracellular matrices independently of collagen. Proc Natl Acad Sci. 1984; 81: 471–475.
- Saelman EUM, Nieuwenhuis HK, Hese KM, de Groot PG, Heijnen HFG, Sage H, et al. Platelet Adhesion to Collagen Types I Through VI11 Under Conditions of Stasis and Flow Is Mediated by GPIa/IIa (a2βl-Integrin). βlood. 1994; 83: 1244-1250.
- Savage B, Almus-Jacobs F, Ruggeri ZM. Specific synergy of multiple substrate–receptor interactions in platelet thrombus formation under flow. Cell. 1998; 94: 657–666.
- Sixma JJ, Groot PG. Regulation of platelet adhesion to the vessel wall. Ann N Y Acad Sci. 1994; 714: 190–199.
- Tokarev AA, Butylin AA, Ataullakhanov FI. Platelet adhesion from shear blood flow is controlled by near-wall rebounding collisions with erythrocytes. Biophys J. 2011; 100: 799–808.
- Caro CG, Fitz-Gerald JM, Schroter RC. Atheroma and arterial wall shear observation, correlation and proposal of a shear dependent mass transfer mechanism for atherogenesis. Proc R Soc London B Biol Sci. 1971; 177: 109–133.
- Fogelson AL, Neeves KB. Fluid Mechanics of Blood Clot Formation. Annu Rev Fluid Mech. 2015; 47: 377–403.
- Wang W, Diacovo TG, Chen J, Freund JB, King MR. Simulation of platelet, thrombus and erythrocyte hydrodynamic interactions in a 3D arteriole with in vivo comparison. PLoS One. 2013; 8: 1-11.
- Vahidkhah K, Diamond SL, Bagchi P. Platelet dynamics in three-dimensional simulation of whole blood. Biophys J. 2014; 106: 2529–2540.
- Casa LDC, Deaton DH, Ku DN. Role of high shear rate in thrombosis. J Vasc Surg. 2015; 61: 1068–1080.
- Turitto VT, Baumgartner HR. Platelet interaction with subendothelium in flowing rabbit blood: effect of blood shear rate. Microvasc Res. 1979; 17: 38–54.
- Bark DL, Para AN, Ku DN. Correlation of thrombosis growth rate to pathological wall shear rate during platelet accumulation. Biotechnol Bioeng. 2012; 109: 2642–2650.
- Mehrabadi M, Casa LDC, Aidun CK, Ku DN. A Predictive Model of High Shear Thrombus Growth. Ann Biomed Eng. 2016; 44: 2339-2350.
- Herbig BA, Diamond SL. Pathological von Willebrand factor fibers resist tissue plasminogen activator and ADAMTS13 while promoting the contact pathway and shear-induced platelet activation. J Thromb Haemost. 2015; 13: 1699–1708.
- Bernardo A, Ball C, Nolasco L, Choi H, Moake JL, Dong JF. Platelets adhered to endothelial cell-bound ultra-large von Willebrand factor strings support leukocyte tethering and rolling under high shear stress. J Thromb Haemost. 2005; 3: 562–570.
- Ruggeri ZM, Orje JN, Habermann R, Federici AB, Reininger AJ. Activation-independent platelet adhesion and aggregation under elevated shear stress. Blood. 2006; 108: 1903–1910.
- Fredrickson BJ, Dong JF, McIntire L V, López JA. Shear-dependent rolling on von Willebrand factor of mammalian cells expressing the platelet glycoprotein Ib-IX-V complex. Blood. 1998; 92: 3684–3693.
- Colace T V, Diamond SL. Direct observation of von Willebrand factor elongation and fiber formation on collagen during acute whole blood exposure to pathological flow. Arterioscler Thromb Vasc Biol. 2013; 33: 105–113.
- Inauen W, Baumgartner HR, Bombeli T, Haeberli A, Straub PW. Dose-and shear rate-dependent effects of heparin on thrombogenesis induced by rabbit aorta subendothelium exposed to flowing human blood. Arterioscler Thromb Vasc Biol. 1990; 10: 607–615.
- Moake JL, Turner NA, Stathopoulos NA, Nolasco LH, Hellums JD. Involvement of large plasma von Willebrand factor (vWF) multimers and unusually large vWF forms derived from endothelial cells in shear stress-induced platelet aggregation. J Clin Invest. 1986; 78: 1456-1461.
- Behroozinia P, Taheri S, Mirzaeifar R. An Investigation of Intelligent Tires Using Multiscale Modeling of Cord-Rubber Composites. Mech Based Des Struct Mach. 2017; 46: 1-16.
- Behroozinia P, Bayandor J, Mirzaeifar R. Numerical Investigation of Scale Factor in Composites Applying Extended Finite Element Method. AIAA Modeling and Simulation Technologies Conference. 2016; 1913.
- Behroozinia P, Mirzaeifar R, Taheri S. A review of fatigue and fracture mechanics with a focus on rubber-based materials. Proc Inst Mech Eng Part L J Mater Des Appl. 2017.
- Nobili M, Sheriff J, Morbiducci U, Redaelli A, Bluestein D. Platelet activation due to hemodynamic shear stresses: damage accumulation model and comparison to in vitro measurements. ASAIO J (American Soc Artif Intern Organs 1992). 2008; 54: 64-72.
- Mokhlespour Esfahani MI, Taghinedjad S, Mottaghitalab V, Narimani R, Parnianpour M, Loughlin C, et al. Novel printed body worn sensor for measuring the human movement orientation. Sens Rev. 2016; 36: 321-331.
- Mokhlespour Esfahani MI, Zobeiri O, Moshiri B, Narimani R, Mehravar M, Rashedi E, et al. Trunk motion system (TMS) using printed body worn sensor (BWS) via data fusion approach. Sensors. 2017; 17: 112-127.
- Mokhlespour MI, Zobeiri O, Akbari A, Milani Y, Narimani R, Moshiri B, et al. Sharif-human movement instrumentation system (SHARIF-HMIS) for daily activities. Biomedical Engineering (ICBME), 2012 19th Iranian Conference of. IEEE. 2012; 143–148.