
Special Article - Malnutrition and Nutritional Deficiency
Austin J Nutri Food Sci. 2018; 6(3): 1106.
Genetically Modified (GM) Crops: A Potential Source to Combat Global Hunger and Malnutrition
Chaudhuri A and Datta A*
National Institute of Plant Genome Research, India
*Corresponding author: Datta A, National Institute of Plant Genome Research, Aruna Asaf Ali Marg, New Delhi- 110067, India
Received: May 25, 2018; Accepted: June 22, 2018; Published: June 29, 2018
Abstract
Genetically modified crops and their use have been controversial since their inception. While pro and anti GM groups are in debate about the consequences unknown to our health and planet’s welfare, it’s now an established fact that GM crops can be a source to combat malnutrition and food shortage not only due to their enhanced yields and nutritional quality but also due to increased resistance to various types of stresses. GM crops are dominant compliments to those produced by conventional plant breeding techniques, which have many drawbacks and are also incompetent to meet the ever increasing demands of the booming global population. To meet these concerns, new advances in crop genetic engineering techniques could be utilized to improve and develop new varieties of food and cash crops. Cutting edge techniques like cis genesis and intra genesis, involving transformation of plants with genetic material borrowed from species itself or from analogous species, with the potential of sexual union respectively are proving to be a boon to enhance nutrition and increased global food production. Modern technologies like genome editing are predicted to go a long way in the future in risk free production and consumption of GM crops. Such advances of GM technology are the only way to combat malnutrition and food scarcity. Need for optimal nutrition and healthy lifestyle holds paramount importance in our lives. Genetic engineering has been largely utilized to enhance nutrition of crops either by reinforcing, by boosting the existing nutrients or by throwing out the toxins or anti nutrients.
Keywords: GM crops; Food production; Nutritional quality; Crop improvement; Shelf life; A biotic Stress; Biotic Stress
Introduction
Genetically modified (GM) crops could be the possible answer to address global hunger. Each year several million children either die or suffer irreparable defects because of malnutrition and starvation. When GM crops were first produced by scientists globally, they were rightly publicized as a big leap forward in the arena of supplementing nature with high nutrient foods. The well fed rich countries of the world probably may not recommend GM foods but the rest of the world badly needed it. According to the United Nations Organization (UNO) survey, it’s recorded that nearly one in eight people across the globe face chronic malnutrition. The discovery of “Golden Rice” where the seeds of rice are fortified with vitamin A precursor betacarotene [1] was breakthrough in GM food research. This discovery was a significant step as a humanitarian gesture which was capable of alleviating the suffering of some 250 million children-the potential cause of preventable blindness and immunodeficiency-including a whopping 2 million dying due to this deficiency.
It was first discovered in 1946 that DNA can be transferred between organisms. But now it’s a well known fact that there are a plethora of mechanisms of DNA transfer from one organism to another in nature on a large scale Figure 1. The first genetically modified crop was produced in 1983 using an antibiotic resistant tobacco and Petunias. The breakthrough technology of DNA transfer with the bacterial pathogen Agrobacterium tumifaciens to plant host was a major contribution by Chilton and his research group [2]. In the early 1990s China took a significant leap by commercializing a transgenic crop with the introduction of virus resistant tobacco. The transgenic “Flavor Savor Tomato” with the trait of delayed ripening was approved in 1994 by the food and Drug Administration (FDA) for marketing in the USA. Other transgenic crops which received market vindication were canola with modified oil composition (Cal gene), Cotton resistant to herbicide Bromoxynil (Cal gene), Bacillius thuringiensis to now (BT) corn (Ciba-Geigy), Bt Cotton (Monsanto), BT potato (Monsanto) and many more. After decades of decry about bio safety of GM crops, concerned authorities cannot now turn around the claim that biotech crops are safe for consumption. To combat the mammoth proportions of the global population especially children who go hungry to bed every night, GM crops could be utilized effectively. This review is an endeavor to compile the already achieved success in enhancement of food production and fortification of nutrition but also the milestones that can be reached through more and more progressive tools and techniques.
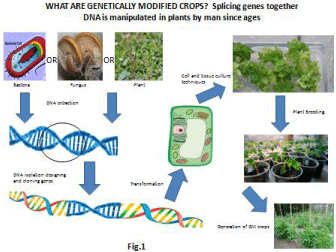
Figure 1: GM technology for genetically improved crop plants with higher yield, nutritional value, enhanced stress tolerance and wider adaptability.
GMOs and GM foods- What and How?
Genetic modification or alteration is a potential biological technique that affects refitting of the genetic machinery of all variety of living organisms. According to the World Health Organization (WHO), GMOs are Organisms (i.e. plants, animals or microorganisms) in which the genetic material (DNA) has been altered through gene transfer technology. With the aid of Recombinant DNA technology, gene transfer can smoothly happen between one organism and another, usually unrelated. The FAO (Food and Agricultural Organization of the United Nations) in association with the European Commission have set clear definitions for GMOs. The GMO definition goes as “not occur naturally by mating and/or natural recombination”. The crux hence lies that GM foods are those which are genetically modified plants or animals. GM crops, our main area of focus are the outcome of very specific and targeted alteration in the plant genome, where the final products such as proteins, metabolites or the phenotypes are well characterized [3]. From another point of view [4] cited the example of Triticale. Triticale is a grain which was developed in the 19th century by the cross of wheat and rye, which turned out to be sterile. In 1930, polyploidy embryo cells were generated using the medicine Colchicines, which were fertile. Thus Triticale was a perfect fit to the definition of GMO and according to Oliver “Biotechnologically Developed organism is a much appropriate definition for GMO.
Need for GM foods
Potential source of nutrition: To cater to the food demands of the booming world population, introduction of a lone gene for development of a single trait will merely suffice. The ever increasing need to develop crops with complex traits such as stress tolerance and nutrient-use efficiency as well as combinations of multiple traits has become the motto of all crop researchers. GM crops have not been conferred as the ‘‘absolute solution’’, it has been established that they could undoubtedly make a noteworthy contribution to an array of measurements and incentives to this invariably growing problem. The forthcoming years will be crucial for the commercial and economically viable application of GMOs in agriculture and food production [5].
Super bananas developed by Professor James Dale with increased level of β-carotene (20 mg per gram dry weight) are under human testing. The phytoene synthesis (PSY2a) gene isolated from the supine banana variety was transformed to create super banana [6]. Most plants have a meager balance of essential amino acids relative to the needs of animals and humans. Betterment of the amino acid composition of the crop plants has been the primary focus of various crop improvement programs. Successful reports of improving essential amino acid like lysine in corn [7], canola and soybean [8] are available. The AmA1 protein, which is a seed albumin from Amaranthus hypochondriacus, is a source of major essential amino acids and is totally safe from the allergen point of view and fits the parameters of optimum nutrition set by the World Health Organization [9]. Work done on transgenic potato expressing AmA1 [10], was a major success where a peak in the growth and production of tubers was observed coupled with a drastic increase the total protein content where all the essential amino acid content showed an increasing trend. Further work on these transgenic potato tubers demonstrated enhanced photosynthetic activity, increase in total biomass, and increase in tuber yield [11]. Multiple desirable traits can be easily introduced in genetically modified crops whereas stacking of traits by conventional breeding is laborious as well as obsolete process. Enhanced drought tolerance, resistance to biotic stress, increased iron and beneficial Polyunsaturated Fatty Acid (PUFA) content has been achieved in tomato by expression of a single fungal gene-C-5 sterol desaturase [12]. This strategy can be further extrapolated to other economically important food and cash crops like rice, soya bean etc.
Genetic engineering can be successfully employed to channelize the secondary metabolites or the Anti Nutrient Factors (ANFs), which are otherwise fatal for human or animal consumption. ANF generation and metabolism can be controlled by the knock down of gene expression associated with their metabolic cycling and recycling. This simple organic acid (Oxalic acid) is a precursor of β-N-oxalyl- L-a, b-diaminopropionic acid (β-ODAP), a deadly neurotoxin found in grass pea. Nephrolithiasis is the pathological condition caused by the deposition of calcium oxalate crystals in kidneys hence causing severe damage. Edible basidiomycetes fungus Flammulina velutypes is the potential source of Oxalate decarboxylase [13] the oxalate degrading enzyme [14]. Showed that the induction of OXDC transcript was a pH dependent phenomenon and is regulated by an EF hand calmodulin like protein–FvCamLP [15,16] Clearly proved that expression (constitutive/seed specific) of OXDC caused a cutback in Oxalic Acid level in soya bean and grass pea (up to73% and 75% respectively). The enzyme OXDC overpowers other oxalate degrading enzymes in many respects. It is highly substrate specific (oxalate), at a considerably low pH, leads to a one step degradation to formic acid and carbon dioxide minus any cofactor [13].
RNAi technology has been utilized on a broad scale to prepare sense and antisense constructs, which are expressed under seed specific promoters to eliminate plant allergens. A big example of RNA interference can be cited in soybean where the p34 protein (a member of the papa in super family of Cys proteases) is suppressed by RNA interference. Similar example can be cited in rice where 14kD to 16kD allergenic proteins have been silenced by antisense.
Stress tolerance: The model plant, Arabidopsis is a well known template for all the experimentations regarding stress perception, sensitivity and tolerance. Deductions got from Arabidopsis have superimposed on a plethora of commercially important food crops such as rice soybean and maize [17]. Elucidated a list of a biotic stresses (salinity, drought and high temperature) which negatively affect the growth, development and ultimately the productivity of crops. Reactive Oxygen Species (ROS) are released on the perception of any type of stress which alters the homeostasis and metabolism of a plant body. AP2/EREBP, Myc, NAC, HB, HSF, bZIP Cys2His2 zincfinger and WRKY are the transcription factors which are activated when stress is perceived by the plant body. These in turn activate distinct stress genes which prepare the plant for defense.
Epicuticular wax forms the first layer of defense against any form of stress. This wax layering on the outermost strata of the aerial plant organs is the protective barrier against insect pests, fungal pathogens as well as drought. This cuticular wax coat can be tailored by working on the gene(s) governing wax metabolic pathway. In fact this is amongst one of the successfully tried and tested techniques to enhance drought tolerance, and thus minimize transpirational water loss. A breakthrough research work was done by [12], in which C-5, sterol desaturase (FvC5SD) was expressed in tomato plants isolated from the fungus Flammulina velutypes. It was observed that there was a pronounced increase in the deposition of epicuticular wax. This phenotypic modification bestowed enhanced drought resistance to the transgenic. As tomato is the natural host of Sclerotinia sclerotiorum, the potential of the fungus to infect the leaves of transgenic plants and wild type plants were also tested. The transgenic lines showed slow progress of the disease compared to the wild type plants because of the thicker wax layer on the outer leaf surface.
Draught resistance can be enhanced in multiple ways especially by refitting the Late Embryogenesis Abundant (LEA) genes in plants. Transgenic Arabidopsis plants are a prominent example. They ectopically express DHN-5, a wheat dehydrin. The characters expressed post gene expression were improved seed germination rate, water retention, growth, with an increased pro line as compared to WT plants put under salt and/or drought stress [18]. Other multiple targets of gene manipulation for drought tolerance achievement are aquaporins and ion transporters, plant helices, compatible solutes like GlyBet [19], ROS scavenging system [20,21], glyoxalase [22], and heat shock proteins [23].
Disease resistance is one of the major traits of GM crops. This trait has been utilized successfully to combat a wide range of fungal and bacterial pathogens without affecting beneficial attributes. The oxalate degrading enzyme, OXDC, expressed in tobacco, lathyrus, tomato, and soybean led to a drastic increase in resisting the pathogen Sclerotinia sclerotiorum that utilizes oxalic acid during its host colonization [16,24]. The defense system controlling machinery in crop plants can be boosted up against various pathogen attack (virus, bacteria, fungi, nematodes, and insects) using RNAi interference strategy [25]. The viral coat protein has been manipulated to mediate resistance to viruses and this has been one of the most successful approaches employed in plant genetic engineering. Many virus resistant plants have been brought to market like PYV tolerant varieties of potato (Potato Y Virus) or PLRV (Potato Leaf Roll Virus) [26]. Reported the generation of transgenic tobacco which expressed the defective Cucumber Mosaic Virus (CMV) replicas’-derived dsRNA, which conferred high resistance to the disease. Resistance against virus infection was also got by utilizing the sense and antisense RNA steering the replication associated protein (AC1) of African Cassava Mosaic or the C1 gene from the Gemini virus. Expressing the defective viral Movement Protein (MP) could also confer resistance to virus [27].
One of the novel approaches to generate virus resistant plants is Post Transcriptional Gene Silencing (PTGS) which generates plants resistant to a large range of viruses. Several examples can be cited where PTGS technique is utilized to confer resistance against DNA viruses which includes Gemini viruses in plants [28]. Have designed a gene silencing vector using the features of transacting small interfering RNA (tasiRNA). They targeted two RNAi suppressor proteins (AC2 and AC4) of the Gemini virus Tomato Leaf Curl New Delhi Virus (ToLCNDV) and successfully developed transgenic tobacco plants with resistance against this important plant virus. Resistance conferred to mixed virus infections can be achieved through multimiRNA strategy [29].
Augmented shelf life: Fruit softening (climacteric and nonclimacteric) during the ripening process is a very delicate process since fruits can be easily damaged by excessive softening thus leading to spoilage and severe loss in both non-climacteric and climacteric fruits. Ethylene is the key hormone governing fruit ripening by the activation of a transcriptional signaling cascade, which regulates and triggers the expression of a plethora genes associated with the fruit ripening process. Biotechnological strategies like RNA interference have been utilized to delay the ripening and increase the shelf life. The ACS and ACO gene are the two potential candidates in the ethylene biosynthesis and action cascade. They have been manipulated by several techniques to delay the ripening and softening process. One such technique was the suppression of genes of ACS (1-aminocyclopropane-1-carboxylicacid synthesis) or ACO (ACC oxidize) [30]. Transforming plants with ACS or ACO gene in antisense direction is one of the techniques adopted to increase the shelf life of fruits [31].
Cell wall degradation during fruit softening is a significant process and the enzyme Polygalacturonase enzyme (PG) plays a very important role modulating the entire process. Antisense/RNAi strategy has been applied to refit to reduce the action of pectin polymerization and make the fruits firm and long lasting [32] keeping the fruit characteristics (weight color, or soluble solids) intact. The ripening process of non-climacteric fruits on the other hand is independent of ethylene. So refitting genes involved in the ethylene biosynthesis or perception pathway can’t be utilized. Hence it becomes imperative to develop a common method of gene reorganization, which can be applied to both climacteric and non climacteric fruits. A significant research work done by [33] suppressed a-mannosidase (a-Man), and β-D-N-acetylhexosaminidase (ß-Hex), N-glycan processing enzymes. These two enzymes enhanced fruit shelf life in tomato by reducing the rate of softening. The shelf life of the transgenic tomato was increased by ≈30 days and there was a ≈ 2.5 fold firmer fruits in a-Man RNAi line and ≈2-fold firmer fruits in case of β-Hex RNAi line. Another breakthrough work by [34] where the nullification of a-Man and β-Hex (RNAi mediated) in capsicum, a non- climacteric fruit, delayed the fruit deterioration by ~7 days. The a-Man and β-Hex RNAi line of fruits were found to be ~2 times firmer with respect to the control fruits. Thus the N-glycan processing enzymes are the potential modulators that can be manipulated to minimize post-harvest losses both for climacteric and non-climacteric fruits. RNAi silenced a-Man and ß-Hex lines, blocks not only degeneration of the N-glycoproteins but also the release of N-glycan required during ripening. The high level of activities in fruits like tomato, suggest their potential involvement in both climacteric and non-climacteric fruits. So it’s a smooth and clear conclusion that, genetic manipulation of N-glycan processing can be applied to modulate fruit ripening in a big and effective way. [35,36] has identified and characterized promoters of β-Hex and a-Man. This research heralded a few novel concepts on the transcriptional regulation of the above genes during fruit ripening. Thus, these specific promoters involved in fruit ripening could potentially useful tools in modulating gene expression during the entire ripening process. Graphic representations of all the above advantages are in Figure 2.
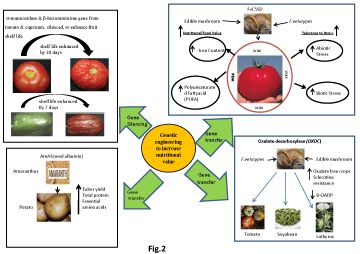
Figure 2: Various approaches for crop improvement.
Emerging tools and techniques in crop genetic engineering
Severe limitations accrue when crop transformation is performed with traditional methods (Agrobacterium-mediated, particle bombardment). The antibiotic resistance gene, which is used as a selection marker in the production of GM crops are a potential risk and have consumer concern factor too, since they escape out in the environment (non-GM crops/related wild species). An upcoming technology- “Site-Specific Recombination” can be utilized to overcome a number of technical difficulties faced during genetic engineering like expression or manipulation of multiple genes in plant genome. Cre-lox from Escherichia coli bacteriophage P1, FLPFRT from S. cerevisiae and R-RS from Zygosaccharomyces rouxi are the three effective SSR systems proposed in 1990s and are still being used till date [37]. Recombines’-mediated excision has also been used in many crop plants like wheat and rice. Marker free transgenic rice and tobacco has been developed using the Transposon based MAT (multi-auto-transformation) vector system. This process involves an oxidative stress-inducible FLP/FRT system to knock out a selectable marker gene [38]. Another strategy is co-transformation to produce marker-free transgenic plants. In this process, co-transformation of a selectable marker gene and a gene of interest from different T-DNAs is mediated by Agrobacterium. This is followed by segregation of the genes, subsequently, in the forthcoming sexual progeny [39]. Marker free transgenic plants can also be generated through Non-selected transformation. Here we can skip the usage of marker genes. This strategy was used in developing peanut transgenic plants (marker free) with binary vector pCAMBIA2300 which has no selection marker gene [40]. The absence of selection marker, on one hand has some advantages but on the other hand it acts as a bottleneck during the screening of the transformed plants.
In order to address all bottlenecks brought forward by the anti- GM groups, against transgenics, “cisgenesis” or “cisgenic” has been proposed as an alternative genetic engineering avenue [41]. Cisgenic plants are so generated where one or more genes are isolated from similar or sexually compatible species and genetically modified. On the other hand an intragene is not a perfect copy of a natural gene and therefore “intragenesis differs from “cisgenesis” in the composition and arrangement of the genetic construct.
Pointed out that these approaches and applications are not only efficient and environment friendly methods for generation of transgenic crops but also they are consumer friendly due to the absence of the vector-backbone sequence and the selectable marker gene [42]. The procedure by which the transgenics are developed by “intragenesis” and “cisgenesis” matches more or less to those generated by traditional breeding as both methods utilize identical gene pool. Whole genome sequencing will prove to be boon for the techniques “intragenesis” and “cisgenesis” in a way that will allow the increased availability of the number of genes and also expand the possibilities for identify paralogues of a gene.
Sequence Specific Nucleases (SSNs) are taking the world by storm and are catergorised as meganucleases, Zinc Finger Nucleases (ZFN) and clustered regularly interspersed short palindromic repeats (CRISPR)/Cas9 reagents and Transcription Activator-Like Effecter Nucleases (TALENs). The utility of Mega nucleases are multifarious although associated with a few bottlenecks. TALENs have come up as the choicest reagents for numerous genome engineering applications. They too, like ZFNs are chimeric proteins generated by soldering an engineered DNA binding region with the catalytic region of Fok1 end nuclease which cuts as a dimmer. The main theme of working of both TALENs and ZFNs are almost same, both work in pairs. The basic advantage of TALENs lie in the fact that DNA binding domain can be engineered smoothly and easily and in the process it can identify virtually any random DNA sequence [43].
The debut and the gradual evolution of the CRISPR/Cas9 technology outmode TALENs and ZFNs and are applied widely not only as a novel but as a breakthrough approach for their super efficient genome editing. This system, in bacteria, is engaged in the defense system of the host from the attack of the viral (phage) and other species of plasmid DNA. This system is rapidly taking the centre stage in the domain of targeted mutagenesis and gene editing in crops, both food and cash crops, because of lack of difficulty in its assembly and considerably greater frequency of bi-allelic mutation. Last five years witnessed many reports highlighting the application of CRISPR/Cas9-based genome editing in plants. Of late, reports on soya bean (Glycine max) and crop have spot lighted the use this breakthrough technology using hairy-root composite plants, soybean protoplasts and whole soya bean plants [44]. And [45] have reported the generation of Gemini virus-resistant Nicotiana benthamiana and Arabidopsis thaliana. Here the Cas9/sgRNA expressed in plant cells successfully interfered with viral replication., Cyanides, glucosinolates, phytate, oxalic acid, neurotoxin, β-N-oxalyl-aminoalanine- l-alanine (BOAA) etc are the common anti- nutrients found in plants which make them unsafe and unfit for human consumption. Genome editing by CRISPR/Cas9, and CRISPR/Cas9 based gene knockouts can alter/modify these genes/biochemical pathways responsible for the biosynthesis of these harmful metabolites. The CRISPR/Cas9 technology can be put to use to improve the multifarious attributes of fruit, thus improving their edibility and shelf life.
Concluding remarks and future perspectives
GM crops, a recent phenomenon, and contains in itself a plethora of unanswered questions. Insertions of genes into other genomes might result in unexpected aftermath. But gradually the concept is seeping in the minds of common man that the genesis of GM crops is truly intended for “public good” rather than corporate profit. In spite of the big debate between the pro and anti GM groups all over the world, GM crops have far more advantages than shortcomings. The introduction of transgenic wheat, that still awaits approval, would cut the wheat exports by 25-50% in the U.S. this in turn could reduce the prices up to 33%. The increasing popularity of genetically engineered DNA/Recombinant DNA (rDNA) techniques prompted mass gathering and discussions regarding public safety in the 1970s. Later on in 1980s it was decided that “DNA was DNA, no matter what the source”.
With a humongous number people in the globe, undernourished, and close to 2 billion of the world’s population suffers from “hidden hunger” [46]. Vitamins and minerals are generally added to the food crops through the practices like supplementation and bio fortification. In conventional breeding the use of fertilizers bio fortified with the desired micronutrients is the most carried out practice. Conventional plant breeding processes also have been so designed and utilized successfully to improve the micronutrient content of plants. This process also has limitations with respect to the extent of variations in the plant gene pool and the optimum time required in producing the desired cultivars with the required traits. Biotechnology has proved as an effective alternative method for the production of micronutrient enriched bio fortified crops (Table 1). This process is cost effective as well as sustainable. Transgenic crop technology enables the gene of interest to be incorporated directly and the resulting transgenic plant expressing the recombinant protein might not have been achievable by conventional breeding. Several benefits have been recorded in case of a few genetically modified crops vis a vis their conventional counterparts [47]. The recent techniques of biotechnology like genome editing have taken the spotlight where the nuclease-based forms of genetic engineering (TALENS, CRISPR, CRISPR associated Cass systems) is the main focus of genome editing. The new generation of genetically engineered food crop with highly beneficial traits like elevated iron storage protein or increased quantities of foliate can replenish the daily diets of the developing world with not only the mentioned nutrients but also with other essential and mandatory micronutrients which generally lack in the diet of the malnourished population [48]. These micronutrients not only should be generated in plants but should also be readily bio available to the human body so that the micronutrient status of the consumer can be improved even after cooking/ processing the food. It’s now a big responsibility on the scientific community to educate and enlighten the farmers and common man so that GM crops are readily adapted and easily accepted by them to increase the general health and nutrition status of the community. At some points of time this factor becomes quite a point of dilemma as some cross-sections of population remain wary about the shortcomings of GM crops. For example in 2002, six African countries refused food aid from the U.S. due to fears of GM presence. With these upcoming skills and powers the basic understanding of a plant’s metabolic cascade can be vividly elucidated and will place the plant researchers working on GM crops on a much firm platform where they can boast the generation of nutritionally enhanced minor and major crops thus improving various aspects of overall well-being.
![]()
CROP
GROSS TRAIT
TRAIT IMPROVED
Maize
Protein level and quality, Mineral
Amino acid composition, Protein↑, Phytase ↑, Ferritin ↑
Rice
Protein level and quality, Carbohydrates, Minerals
Amino acid composition, Protein↑, Amylase↑, Iron ↑
Potato
Protein level and quality, Essential amino Acids, Carbohydrates
Amino acid composition, Protein↑, Methionine ↑
Canola
Essential Amino Acids, Vitamins
Lysine ↑, Vitamin E ↑
Sorghum
Essential Amino Acids
Lysine ↑
Lupin
Essential Amino Acids
Methionine ↑
Soybean
Protein level and quality, Carbohydrates
Amino Acid balance; Fructan, Raffinose
Chicory
Carbohydrates
Fructan ↑
Cotton
Oils and Fatty Acids
Oleic+ Stearic Acid ↑, Oleic Acid ↑
Linseed
Oils and Fatty Acids
+?-3 and ?-6 fatty acids
Palm Oil
Oils and Fatty Acids
Oleic/Stearic Acid ↑, Oleic Acid ↑, Palmitic Acid ?
Safflower
Oils and Fatty Acids
GLA ↑
Mustard
Carotenoids
+β-carotene
Tomato
Carotenoids
Folate ↑, Phytoene and β-Carotene ↑, Lycopene ↑, Provitamin A ↑
Strawberry
Vitamins
Vitamin C ↑
Apple
Functional Secondary Metabolite
+ stilbenes
Alfalfa
Functional Secondary Metabolite, Minerals
+ resveratrol, Phytase ↑
Lettuce
Mineral
Iron ↑
Table 1: Crops developed by advanced genetic engineering tools.
References
- Ye X, Al-Babili S, Kloti A, Zhang J, Lucca P, Beyer P, et al. Engineering the provitamin A (beta-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science. 2000; 287: 303-305.
- Chilton MD. A vector for introducing new genes into plants. Sci Am. 1983; 248: 36-45.
- Datta A. Genetic engineering for improving quality and productivity of crops. Agriculture & Food Security. 2013; 2: 15.
- Oliver MJ. Why we need GMO crops in agriculture, MO Med. 2014; 111: 492-507.
- Nap JP, Metz PLJ, Escaler M, Conner AJ. The release of genetically modified crops into the environment part I. Plant J. 2003; 33: 1-18.
- Mlalazi B, Welsch R, Namanya P, Khanna H, Geijskes RJ, Harrison MD, et al. Isolation and functional characterization of banana phytoene synthase genes as potential cisgenes. Planta. 2012; 236: 1585-1598.
- Eggeling L, Oberle S, Sahm H. Improved L-lysine yield with Corynebacterium glutamicum: use of dapA resulting in increased flux combined with growth limitation. Appl Microbiol Biotechnol. 1998; 49: 24-30.
- Falco SC, Guida T, Locke M, Mauvais J, Sanders C, Ward RT, et al. Transgenic canola and soybean seeds with increased lysine. Bio/Technology. 1995; 13: 577-582.
- Raina A, Datta A. Molecular cloning of a gene encoding a seed-specific protein with nutritionally balanced amino acid composition from Amaranthus. Proc Natl Acad Sci USA. 1992; 89: 11774-11778.
- Chakraborty S, Chakraborty N, Datta A. Increased nutritive value of transgenic potato by expressing a non allergenic seed albumin gene from Amaranthus hypochondriacus. Proc Natl Acad Sci USA. 2000; 97: 3724-3729.
- Chakraborty S, Chakraborty N, Agrawal L, Ghosh S, Narula K, Shekhar S, et al. Next generation protein rich potato by expressing a seed protein gene AmA1 as a result of proteome rebalancing in transgenic tuber. Proc Natl Acad Sci USA. 2010; 41: 17533-17538.
- Kamthan A, Kamthan M, Azam M, Chakraborty N, Chakraborty S, Datta A. Expression of a fungal sterol desaturase improves tomato drought tolerance, pathogen resistance and nutritional quality. Sci Rep. 2012; 2: 951.
- Mehta A, Datta A. Oxalate decarboxylase from Collybia velutipes: Purification, characterization and cDNA cloning. J Biol Chem. 1991; 266: 23548-23553.
- Azam M, Kesarwani M, Chakraborty S, Natarajan K, Datta A. Cloning and characterization of 5'-flanking region of oxalate decarboxylase gene from Flammulina velutipes. Biochem J. 2002; 367: 67-75.
- Kamthan A, Kamthan M, Kumar A, Sharma P, Ansari S, Thakur SS, et al. A Calmodulin like EF hand protein positively regulates oxalate decarboxylase expression by interacting with E-box elements of the promoter. Sci Rep. 2015; 5: 14578.
- Kumar V, Chattopadhyay A, Ghosh S, Irfan M, Chakraborty N, Chakraborty S, et al. Improving nutritional quality and fungal tolerance in soyabean and grass pea by expressing an oxalate decarboxylase. Plant Biotechnology Journal. 2016; 14: 1394-1405.
- Varshney RK, Bansal KC, Aggarwal PK, Datta SK, Craufurd PQ. Agricultural biotechnology for crop improvement in a variable climate: hope or hype?. Trends Plant Sci. 2011; 16: 363-371.
- Brini F, Yamamoto A, Jlaiel L, Takeda S, Hobo T, Dinh HQ, et al. Pleiotropic effects of the wheat dehydrin DHN-5 on stress responses in Arabidopsis. Plant Cell Physiol. 2011; 52: 676-688.
- Quan R, Shang M, Zhang H, Zhao Y, Zhang J. Engineering of enhanced glycine betaine synthesis improves drought tolerance in maize. Plant Biotechnol J. 2004; 2: 477-486.
- Faize M, Burgos L, Faize L, Piqueras A, Nicolas E, Barba-Espin G, et al. Involvement of cytosolic ascorbate peroxidase and Cu/Zn-superoxide dismutase for improved tolerance against drought stress. J Exp Bot. 2011; 62: 2599-2613.
- Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem. 2010; 48: 909-930.
- Singla-Pareek SL, Reddy MK, Sopory SK. Genetic engineering of the glyoxalase pathway in tobacco leads to enhanced salinity tolerance. Proc Natl Acad Sci USA. 2003; 100:14672-14677.
- Chauhan H, Khurana N, Nijhavan A, Khurana JP, Khurana P. The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant Cell Environ. 2012; 35: 1912-1931.
- Kesarwani M, Azam M, Natarajan K, Mehta A, Datta A. Oxalate decarboxylase from Collybia velutips: molecular cloning and its over expression to confer resistance to fungal infection in transgenic tobacco and tomato. J Biol Chem. 2000; 275: 7230-7238.
- Kamthan A, Chaudhuri A, Kamthan M, Datta A. Small RNAs in plants: recent development and application for crop improvement. Front Plant Sci. 2015; 6: 208.
- Otang NV, Kynet K, Khan RS, Ohara M, Goto Y, Wantanabe M, et al. Transgenic tobacco lines expressing defective CMV replicase derived dsRNA are resistant to CMV-O and CMV-Y. Mol Biotechnol. 2014; 56: 50-63.
- Peiró A, Canizares MC, Rubio L, López C, Moriones E, Aramburu J, et al. The movement protein (NSm) of Tomato spotted wilt virus is the avirulence determinant in the tomato Sw5 gene-based resistance. Mol Plant Pathol. 2014; 15: 802-813.
- Singh A, Taneja J, Dasgupta I, Mukherjee SK. Development of plants resistant to tomato Gemini viruses using artificial trans-acting small interfering RNA. Mol Plant Pathol. 2015; 16: 724-734.
- Duan CG, Wang CH, Fang RX, Guo HS. Artificial microRNAs highly accessible to targets confer efficient virus resistance in plants. J Virol. 2008; 82: 11084-11095.
- Hamilton AJ, Baulcombe DC. A species of small antisense RNA in post-transcriptional gene silencing in plants. Science. 1999; 286: 950-952.
- Litz RE, Padilla G. Genetic transformation of fruit trees. Genomics of tree crops. Springer, Berlin. 2012: 117-153.
- Atkinson RG, Sutherland PW, Johnston SL, Gunaseelan K, Hallett IC, Mitra D, et al. Down-regulation of polygalacturonase 1 alters firmness, tensile strength and water loss in apple (Malus domestica) fruit. BMC Plant Biol. 2012; 12: 129.
- Meli VS, Ghosh S, Prabha TN, Chakraborty N, Chakraborty S, Datta A. Enhancement of fruit shelf life by suppressing N-glycan processing enzymes. Proc Natl Acad Sci USA. 2010; 107: 2413-2418.
- Ghosh S, Meli VK, Kumar A, Thakur A, Chakraborty N, Chakraborty S, et al. The N-glycan processing enzymes α-mannosidase and β-D-1 N acetylhexosaminidase are involved in ripening-associated softening in the non climacteric fruits of capsicum. J Exp Bot. 2011; 62: 571-582.
- Irfan M, Ghosh S, Meli VS, Kumar A, Kumar V, Chakraborty N, et al. Fruit Ripening Regulation of α-Mannosidase Expression by the MADS Box Transcription Factor RIPENING INHIBITOR and Ethylene. Front Plant Sci. 2016; 7: 10.
- Kamthan A, Chaudhuri A, Kamthan M, Datta A. Genetically Modified (GM) crops: milestones and new advances in crop improvement. Theor Appl Genet. 2016; 129: 1639-1655.
- Woo HJ, Cho HS, Lim SH, Shin KS, Lee SM, Lee KJ, et al. Auto-excision of selectable marker genes from transgenic tobacco via a stress inducible FLP/FRT site-specific recombination system. Transgenic Res. 2009; 18: 455-465.
- Tuteja N, Verma S, Sahoo, RK, Ravendar S, Reddy IN. Recent advances in development of marker-free transgenic plants: regulation and biosafety concern. J Biosci. 2012; 37: 167-197.
- Bhatnagar M, Prasad K, Mathur PB, Narasu ML, Waliyar F, Sharma KK. An efficient method for the production of marker-free transgenic plants of peanut (Arachis hypogaea L.). Plant Cell Rep. 2010; 29: 495-502.
- Schouten HJ, Jacobsen E. Cisgenesis and intragenesis, sisters in innovative plant breeding. Trends Plant Sci. 2008; 13: 260-261.
- Vanblaere T, Szankowski I, Schaart J, Schouten H, Flachowsky H, Broggini GA, et al. The development of a cisgenic apple plant. J Biotechnol. 2011; 154: 304-311.
- Reyon D, Tsai SQ, Khayter C, Foden JA, Sander JD, Joung JK. FLASH assembly of TALENs for high-throughput genome editing. Nat Biotechnol. 2012; 30: 460-465.
- Baltes NJ, Voytas DF. Enable plant synthetic biology through genome engineering. Trends Biotechnol. 2015; 33: 120-131.
- Ji X, Zhang H, Zhang Y, Wang Y, Gao C. Establishing a CRISPR-Cas-like immune system conferring DNA virus resistance in plants. Nat Plants. 2015; 1: 15144.
- Hefferon KL. Nutritionally enhanced food crops; Progress and Perspectives. Int J Mol Sci. 2015; 16: 3895-3914.
- De Steur H, Blancquaert D, Strobbe S, Lambert W, Gellynck X, van der Straeten D. Status and market potential of transgenic biofortified crops. Nat Biotechnol. 2015; 33: 25-29.
- Sperotto RA, Ricachenevsky FK, Waldow Vde A, Fett JP. Iron biofortification in rice: It’s a long way to the top. Plant Sci. 2012; 190: 24-39.