
Research Article
Austin J Psychiatry Behav Sci. 2015;2(1): 1036.
Genes, Religion, and Response to Religious vs Conventional Psychotherapy: A Randomized Clinical Trial in Medically Ill Patients with Major Depression
Harold G. Koenig*1,2,3,4, Blanca Gutierrez5, Jorge Cervilla5, Michelle J. Pearce1,4,6, Noha Daher7,8, Bruce Nelson9, Sally F. Shaw9, Harvey Jay Cohen2,4,10 and Michael B. King11
¹Department of Psychiatry and Behavioral Sciences, Duke University Medical Center, USA
²Department of Medicine, Duke University Medical Center, USA
³Department of Medicine, King Abdulaziz University, Saudi Arabia
4Center for Spirituality, Theology and Health, Duke University, USA
5Institute of Neuroscience and Department of Psychiatry, School of Medicine, University of Granada, Spain
6School of Medicine, University of Maryland, USA
7Department of Epidemiology, Biostatistics, and Population Medicine, Loma Linda University, USA
8Allied Health Studies, School of Allied Health Professions, Loma Linda University, USA
9Department of Research, Glendale Adventist Medical Center, USA
10Center for the Study of Aging and Human Development, Duke University Medical Center, USA
11Division of Psychiatry, Faculty of Brain Sciences, University College London, UK
*Corresponding author: Harold G. Koenig, Department of Psychiatry and Behavioral Sciences, Duke University Medical Center, Box 3400, Durham, NC 27710, USA
Received: April 08, 2015; Accepted: April 27, 2015; Published: April 30, 2015
Abstract
Objective: We examine associations between religiosity and polymorphisms of the serotonin and monoamine oxidase genes, and the effect of genotype on response to religious cognitive behavioral therapy (RCBT) vs. conventional CBT (CCBT).
Methods: 132 persons with chronic illness and major depressive disorder were recruited into a clinical trial to evaluate the efficacy of RCBT vs. CCBT. Four functional polymorphisms were assessed: 5-HTTLPR and rs25531 at the serotonin transporter gene (SLC6A4), rs6295 at 5-HT1A receptor gene (HTR1A), and uMAOA-VNTR at the monoamine oxidase A gene (MAOA).
Results: Few associations were found between these polymorphisms and religious involvement, although they were consistent. Religious attendance was less frequent in those with one or more high-risk genotype in the overall sample. In post-hoc analyses, Blacks with LG genotypes of the rs25531 polymorphism were less likely to attend religious services or have daily spiritual experiences, and Blacks with high risk alleles of any gene were also less likely to attend religious services. Among men, daily spiritual experiences and overall religiosity were also lower in the presence of any high risk alleles. Genotype had no effect on response to RCBT vs. CCBT in the overall sample, except in participants with the C/C genotype of HTR1A (group x time interaction B=3.33, SE=1.17, t=2.86, p=0.006, d=0.73) and those with no high risk genotype (group x time interaction B=3.40, SE=1.60, t=2.12, p=0.042, d=0.75) who were more likely to respond to RCBT.
Conclusion: Little overall relationship was found between genotype and either religiosity or response to RCBT. Whether high risk genotypes in certain subgroups of depressed persons may be associated with religious involvement or affect treatment response remains unclear.
Keywords: depression, religiosity, polymorphisms, serotonin transporter, monoamine oxidase, 5-HT1A receptor, cognitive behavioral therapy
Introduction
Since publication of the “God Gene” by Hamer in 2004, there has been speculation that genetic predispositions to religious or spiritual experiences may exist [1]. Rowthorn argued that people “who carry a certain ‘religiosity’ gene are more likely than average to become or remain religious [2]. Based on what he calls the dual inheritance model, the increased fertility of religious persons along with high rates of defection from religion are likely to spread religious genes (or at least genes that confer tendencies toward authoritarianism and conservatism) more widely across the general population. Furthermore, an examination of twin studies by Koenig and Bouchard suggests that religiousness may have considerable genetic roots, with genes explaining as much as 40% to 60% of the tendency towards religious involvement [3]. If true, the genetic basis for such predispositions is likely to be far from simple and probably involves a complex interaction between single genes, multiple combinations of genes, personal environment, and even the surrounding culture. For example, Sasaki and colleagues reported that the rs53576 polymorphism at the oxytocin receptor gene interacted with the cultural environment to affect the relationship between religiosity and well-being [4]. They found that among Koreans in Korea with the G/G genotype (thought to confer social sensitivity [5]), religiosity was associated with greater psychological well-being, whereas among Europeans in America with the G/G genotype, religiosity was associated with lower psychological well-being. Researchers hypothesized that religion may benefit those who are more socially sensitive due to a genetic predisposition, but only if the cultural context supports it.
Genetic polymorphisms and depression
Research also suggests that genetic factors are involved in the risk of developing depression, and the capacity for spiritual or religious experience may be linked to such genes, perhaps affecting a religious person’s vulnerability to depression. One of the most likely candidates is the serotonin transporter gene (SLC6A4), the most studied of all genetic factors in psychiatry. The promoter region of SLC6A4 contains a polymorphism (5-HTTLPR) characterized by a short (S) and a long (L) allele. The S allele produces less serotonin transporter mRNA transcription, resulting in less serotonin transporter protein [6]. Less transporter protein means less effective removal of serotonin from the synapse, making it less available for reuse by the presynaptic neuron. There is also a single nucleotide polymorphism (rs25531) located within the inserted fragment of the 5-HTTLPR, which seems to further affect SLC6A4´s transcriptional activity. The rs25531 allele LA has higher levels of mRNA transcription, while the rs25531 allele LG has levels similar to S allele carriers [7].
In 2003, Caspi and colleagues reported that among individuals with the 5-HTTLPR genotype S/S or L/S, number of stressful life events predicted the development of major depression [8]. This effect was absent in those with the L/L genotype. In 2009, this association was challenged by a meta-analysis of existing studies, concluding that the relationship was uncertain [9]. This meta-analysis, however, was itself challenged because of the methodology used, causing critics to conclude that more research was needed on how SLC6A4 allelic variations affect response to environmental stressors [10,11]. While few studies have examined the relationship between 5-HTTLPR polymorphisms and depression in those with chronic medical illness, where health-related stressors abound, the area is a promising one for future research. For example, in a report involving 737 primary care outpatients, depressive episodes were significantly more frequent (50% to 79%) among those with the S/S genotype, an association that increased in strength with increasing severity of depression [12]. The S/S genotype interacted with number of stressful life events (SLE) such that those with this genotype required only minimal exposure to stressful events to significantly increase risk for depression, compared to L/S or L/L genotypes that required much higher levels of stress to increase risk [13]. These reports are supported by research in patients with heart disease that demonstrate greater vulnerability to depression in those with the S allele [14,15]. Likewise, relationships between the low functioning LG variant (rs25531) and depression, anxiety, and neurosis have also been reported [16,17], especially in those with medical illness [18,19].
Such relationships, however, appear to be influenced by race and gender. While 50% of Caucasians have the S-allele, only 30% of Black Americans do [20]. In contrast, while only 8% of Caucasians have the LG allele at rs25531, 21% of Black Americans do [21]. Furthermore, the effect on depression risk (as reflected by low levels of CSF 5-HIAA) may be the opposite in African-Americans where the L allele has been associated with greater risk rather than the S allele [22]. Gender too has an influence. While the prevalence of S allele is similar in men and women, sensitivity to stress may be greater for women with the S allele than for men. For example, one study found that the L/L genotype conferred a greater risk of depression in stressed males (i.e., male caregivers or males with low education fathers), whereas the S/S genotype conferred a greater risk of depression in stressed females [23].
Besides polymorphisms at the promoter region of SLC6A4, other genetic variants are known to increase risk of depression in persons with medical illness, including the high activity alleles of the promoter region of the MAOA gene [24]. Alleles 3.5, 4, and 5 are associated with high MAOA activity, while allele 3 is associated with low MAOA activity [25,26]. High activity alleles have been reported to increase the risk depression [27,28] and have been linked to personality traits such as neuroticism [29]. Likewise, a polymorphism of the 5-HT1A gene (rs6295) that involves a C to G substitution resulting in C/G and G/G genotypes has been linked to major depression and suicidal thoughts [30,31].
Polymorphisms and religion
While religiousness is often inversely related to depression and associated with faster recovery from depression in the medically ill [32,33], some studies suggest that religious persons may actually be more vulnerable to depression [34,35], perhaps due to greater emotional sensitivity (a trait that Freud called “neurosis” [36]). A common genetic predisposition could provide a biological basis for the link between religion and depression, either making religious persons more or less vulnerable to depression.
Why might religiousness/spirituality be associated with genes that increase sensitivity to life events or increase risk of depression? Although some have suggested that depression is adaptive (bolstering immune systems or encouraging people to think more intensely about their problems), depression and suicide in particular may also serve as an evolutionary force that removes “less fit” persons from the population [37,38]. Religious involvement, in turn, may counter this evolutionary force by providing beliefs that facilitate coping with negative life events by surrounding the depressed individual with a supportive faith community, enabling depressed religious persons to survive and pass on their genes to the next generation. Thus, genes associated with emotional sensitivity to negative life events (increasing the risk of depression) and those associated with religious involvement may be preserved within the population.
Alternatively, perhaps religious persons have genes that somehow make them less prone to depression, although how such genes might have developed and persisted in the population is not clear. However, this would help to explain the inverse relationship between religiosity and depression reported in the majority of studies examining this association [39]. Although such notions are highly speculative, they provide a plausible reason for why a genetic link might exist.
Some research appears to substantiate a relationship between polymorphisms of the SLC6A4 gene and religious involvement, although the findings are not very consistent. For example, the S allele of the 5-HTTLPR polymorphism has been associated with high scores on the Tellegen Absorption Scale (TAS), which assesses the tendency to have mystical visionary experiences or artistic sensitivity [40]. The TAS, in turn, has been correlated with high levels of intrinsic religiosity [41]. These findings are consistent with the claim that the S allele conveys both greater emotional sensitivity and greater sensitivity to spiritual experiences as well. Likewise, the S allele has been associated with higher scores on the spiritual acceptance subscale of the TCI [42] and with other factors that may be considered broadly spiritual [43].
We found no studies that examined relationships between spiritual or religious involvement and 5-HTTLPR-rs25531 polymorphisms (at the serotonin transporter gene) or uMAOA-VNTR (at the MAOA gene). However, a polymorphism (rs6295) of the HTR1A gene (that affects the binding potential of 5-HT1A neuroreceptor for serotonin involving a C to G substitution resulting in C/G and G/G genotypes) has been linked to high scores on the TCI’s spiritual acceptance scale [44]. This finding is consistent with results from functional magnetic resonance imaging that indicate lower brain serotonin receptor binding in those with high scores on the spiritual acceptance scale, suggesting greater vulnerability to depression [45].
Especially unclear is how functional variability at SLC6A4 (or other candidate genes that increase depression risk) moderate response to religious/spiritual interventions. The S and LG alleles have not only been reported to cause greater emotional sensitivity to environmental stressors, but the presence of these low functioning alleles has also been shown to decrease response to treatment in those with major depression [46,47,48]. While the presence of these alleles in those who are more religious could increase vulnerability to depression, greater distress in response to stressors and failure to respond to conventional treatments could also cause individuals to turn to religion for comfort. Since cognitive defects (such as greater negative schematic processing) following negative life events have been documented in those with the S allele [49], perhaps a psychological treatment that utilizes religious resources to counteract negative cognitive schematic processing resulting from chronic medical illness may be particularly effective in those with this form of the gene.
Thus, more research is needed to determine whether a relationship exists between religious involvement and 5-HTTLPR or other neurotransmitter-related genotypes, and how effective religious interventions might be in those with genetic polymorphisms that increase depression risk. To our knowledge, no study has yet examined the relationships between religiosity and high risk genotypes of the 5-HTTLPR (S/S), 5-HTTLPR-rs25531 (LG), HTR1A (G/G), or uMAOA-VNTR (high activity) polymorphisms in depressed persons with chronic medical illness. Likewise, no study has examined how the presence of these polymorphisms affects response of major depression to cognitive behavioral therapy (CBT) (either conventional or religious). Religious CBT, which involves the integration of religious beliefs of patients into psychotherapy to combat dysfunctional cognitions and discourage depressioninducing behaviors (e.g., social isolation) [50], has been shown to be effective in patients with major depression and chronic medical illness [51].
Hypotheses
Using a candidate gene approach, we sought to identify genetic polymorphisms related to religiosity and depression in medically ill patients. We hypothesize that (1) higher religiosity will be positively related to genotypes that place individuals at higher risk for depression, i.e., S/S of the 5-HTTLPR polymorphism, LG of the 5-HTTLPR-rs25531, G/G of HTR1A-rs6295 polymorphism, and presence of any high activity alleles (3.5, 4, or 5) of the uMAOAVNTR; (2) that the effectiveness of RCBT will be greater than CCBT in relieving depression in persons with these genotypes; and (3) the effect of RCBT will be especially strong in those who are highly religious with these genotypes.
Methods
Study design and participants
Participants were enrolled in a two-site randomized clinical trial (Durham, North Carolina, and Glendale, California) to compare the efficacy of RCBT and CCBT in the treatment of major depression. Inclusion criteria were (1) age 18 to 85; (2) one or more chronic medical condition; (3) an affirmative response to the question “Is religion/spirituality at least somewhat important in your daily life?”; (4) a DSM-IV diagnosis of major depressive disorder, and (5) moderately severe depressive symptoms (a score of 10-40 on the Beck Depression Inventory – see below). Exclusion criteria were (1) significant cognitive impairment based on a score of 13 or lower on the abbreviated Mini-Mental State Exam (range 0-18) [52]; (2) receipt of psychotherapy in past two months; (3) psychotic disorder, alcohol or substance abuse, or PTSD within the past year, or history of bipolar disorder; (4) active suicidal thoughts; and (5) diagnosis of HIV/AIDS, autoimmune diseases, dementia, endocrine disorders affecting stress hormone levels, or taking immuno-suppressant drugs (due to other planned analyses). The Duke University Medical Center institutional review board and Glendale Adventist Medical Center approved the study.
Measures
The MINI Neuropsychiatric Inventory [53] was used to diagnose major depressive disorder and to rule out diagnoses that excluded participants from the study (including active suicidal thoughts). The Beck Depression Inventory (BDI) [54] assessed symptom severity (scores of 10 to 40) for inclusion into the study and for tracking symptom severity in response to treatment. Among measures of physical health was the Duke Activity Status Index [55] that assessed physical functioning and the Cumulative Illness Rating Scale [56] that assessed severity of medical illness. Social support was measured using the social interaction and subjective support subscales of the Duke Social Support Index [57]. Religious involvement was assessed with single item measures of importance of religion, religious attendance, and private religious activity, and with multi-item scales assessing intrinsic religious motivation (Hoge IR Scale [58]), religious/spiritual experiences (Daily Spiritual Experiences Scale [59]), and religious coping (Brief RCOPE [60]). In order to increase the power for analysis of the primary hypotheses, the religious variables above were summed to create an overall religiosity measure that ranged from 44- 153 (Cronbach’s alpha=0.88 standardized).
Procedure
Study coordinators screened potential participants by telephone for eligibility criteria, and then arranged a visit when full written informed consent was obtained and an in-person screening evaluation was conducted. Eligible clients were enrolled in the trial, completed a baseline evaluation, and had their blood drawn for genotype analyses. Participants were then randomized to either RCBT or CCBT (see elsewhere for full study details [51,61]). Study coordinators who conducted the screening, baseline, and follow-up evaluations were blinded to treatment group.
The intervention in both groups consisted of ten 50-minute sessions, administered by master’s degree level therapists and delivered over 12 weeks. Sessions were delivered remotely by telephone (90%), Skype (7%), or online (3%) to make it easier for those with physical disability to receive the therapy. CCBT was a manual-based intervention following CBT as described by Aaron and Judith Beck [62,63]. The four therapists who delivered CCBT were experienced in CBT, but had no experience integrating religious beliefs into therapy and if religious issues came up, addressed them in the broadest conventional way possible.
The four therapists who delivered RCBT were experienced with integrating religious beliefs into CBT. RCBT was a manual-based intervention specific to the particular religion of the client [50].CCBT and RCBT interventions were designed to be similar in all respects except that RCBT utilized clients’ religious beliefs in therapy to motivate change in cognition and behavior, whereas CCBT did not. The primary endpoint for the clinical trial was BDI score, which was assessed at baseline, 4 weeks, 8 weeks, 12 weeks (immediately after treatment ended), and 24 weeks from baseline. Individual trajectories of response on the BDI from baseline to the primary endpoint at 12 weeks were determined for participants categorized by genotype.
Blood collection, DNA isolation and genotyping
A 3-4 milliliter whole blood sample was collected in an EDTA tube, followed by mixing well to avoid small clots from forming. Blood samples were stored at –80°C prior to shipping to the Fragment Analysis Facility, Johns Hopkins University, Baltimore MD, where genotype analyses were done. The DNA was isolated from blood using an established interface with the PUREGENE DNA purification system (Gentra systems, Minnesota, USA). The PUREGENE DNA Isolation Kit is a gentle DNA Isolation procedure that isolates DNA using detergents and salts. The PUREGENE DNA Isolation Kit does not contain any toxic chemicals and isolates DNA in high yield from a variety of sample sources. Briefly, the PUREGENE Kit isolates DNA by first using a hypotonic Red Blood Cell Lysis Solution to selectively lyse red blood cells. Then, the Cell Lysis Solution lyses the white blood cells. RNA is then removed if necessary using the RNase A Solution provided in the Kit, and proteins are precipitated and removed using the Protein Precipitation Solution. The DNA was then precipitated out with alcohol, washed, dried and re-hydrated. Analyses were conducted for the 5-HTTLPR and rs25531 at the serotonin transporter gene, the rs6295 at the 5-HT1A receptor gene (HTR1A), and the uMAOA-VNTR polymorphism at the MAOA gene using standardized molecular procedures based on PCR reactions and conventional electrophoresis techniques.
The serotonin transporter (5-HTTLPR) short (S) and long (L) alleles were PCR amplified using forward primer 5’-TCTCCCGCCTGGCGTTGC-3’ and reverse primer 5’-GCCGGTTGGGCTGAGCGTCT-3’. PCR was performed in a 10μl reaction consisting of 0.4μM primers, 0.15μM 7-deaza dGTP, 1X MasterAmp™ 2X PCR PreMix K (Epicenter Biotechnologies, Madison, WI), 1 unit Taq DNA polymerase (Applied Biosystems, Foster City, CA), and 40 ng DNA. Amplification was carried out in a Veriti thermocycler (Applied Biosystems) for 4 min at 95°C, followed by 35 cycles of 94°C for 30 s, 62.8°C for 30 s, and 72°C for 30 s, followed by a final extension step of 72°C for 10 min. PCR products were resolved on 2% NuSieve (FMC BioProducts, Vallensbaek, Denmark), 1% agarose (Invitrogen, Carlsbad, CA) gels, and visualized by ethidium bromide staining. The short and long alleles were characterized by 515 bp and 560 bp products, respectively.
Genotyping of the 5-HTTLPR polymorphism (rs25531) was performed using primers and probes described by Hu et al [64]. PCR was carried out in 5μl volume with 20ng DNA, 0.48μM primers, 0.48μM VIC labeled G allele probe, 0.2μM FAM labeled A allele probe, 1X Master Mix (Applied Biosystems, Foster City, CA), and 4% DMSO (Sigma). Amplification and analysis was performed on an Applied Biosystems 7900HT Sequence Detection System. PCR conditions were 2 min at 50°C, 10 min at 50°C, followed by 45 cycles at 96°C for 30 sec and 66°C for 60 sec. Genotypess were determined using end-point analysis with SDS v2.1 software (Applied Biosystems). We classified the 5-HTTLPR (rs25531) polymorphism into three types: (1) LA/LA, (2) LA/LG, and (3) LG/LG.
Primer sequences for the MAOA uVNTR are MAO APT1 (5’- ACAGCCTGACCGTGGAGAAG-3’) and MAO APB1 5’-ACAGCCTGACCGTGGAGAAG-3’ described by Sabol et al. [12]. The MAO APT1 was 5’-labeled with 6FAM fluorophore. PCR was carried out in 10ul containing 0.1uM primers, 0.1mM dNTPs (Amersham), 10mM Tris (pH8.3); 50mM KCL, 1.5mM MgCl, 0.6 units of Taq DNA polymerase (Perkin Elmer) and 40ng DNA. Amplification was carried out in a Thermo Hybaid MBS 0.2S (Needham Heights, MA) using the following cycling conditions: initial 4-min denaturing step at 94°C, followed by 35 cycles of 94°C for 30 sec., 58°C for 30 sec. and 72°C for 30 sec., followed by a final extension of 72°C for 10 min. PCR products were assayed on a 3730 DNA Analyzer (Applied Biosystems). Data were collected and analyzed with GeneMapper software (Applied Biosystems) that calculated fragment length in reference to an internal lane standard (Genescan-500 labeled with LIZ) and quantitated the amount of fluorescence in each fragment.
Genotyping of the 5-HT1A gene rs6295 polymorphism was carried out using a pre-designed TaqMan® SNP Genotyping Assay (Applied Biosystems, Foster City, CA) following manufacturers supplied protocols. PCR and endpoint detection of fluorescence was carried out in an ABI Prism7900HT Sequence Detection System (Applied Biosystems, Foster City, CA) using default settings. Fluorescence data was analyzed with ABI Prism 7900 allelic discrimination software.
Statistical Analysis
Descriptive statistics were used to characterize the overall sample and assess the comparability of the randomized groups at baseline. analysis of variance (ANOVA) or Student’s t-test was used to determine if religious activities (attendance at services, private prayer or scripture reading), intrinsic religiosity, daily religious/spiritual experiences, religious coping, or overall religiosity varied by genotype. Given known effects of race and gender on the polymorphisms examined here, analyses were stratified by these demographics in post-hoc analyses. Growth curve modeling using random intercept and slope (mixed effect regression models) was used to compare individual trajectories of response to RCBT vs. CCBT in participants with each genotype and in those with any high risk genotype (S/S, LG, G/G, and/or high activity MAOA-uVNTR alleles 3.5, 4, or 5). This was done for the overall sample and the sample dichotomized at the mid-point based on the overall religiosity score (low vs. high). Given the exploratory nature of these analyses, the significance level was set at p<0.05 and was not corrected for multiple comparisons. Statistical analyses were carried out using SAS (version 9.3; SAS Institute Inc., Cary, North Carolina).
Results
A total of 132 participants were recruited at baseline into the trial. Mean age was 51.6 years, average education was 15.1 years, and the majority was female (68.9%). The racial composition of the sample was Caucasian (53.0%), Hispanic (24.2%), Black (16.7%), Asian (5.3%), and other (0.8%). The distribution of 5-HTTLPR, rs25531, rs6295, and uMAOA-VNTR high risk genotypes for depression/ neurosis was as follows: S/S (21.9%), LG (21.3%), G/G (21.1%), and any MAOA high activity allele (3.5, 4 or 5) (74.4%). With regard to the clinical trial, there were no significant differences on baseline characteristics between participants receiving CCBT vs. RCBT, except for the 5-HTTLPR allele distribution (Table 1). No association was found between depression severity and overall religiosity or between depression severity and genotype at baseline (analyses not shown).
![]()
Overall Sample
(n=67)
Conventional CBT
(n= 65)
Religious CBT
(n= 65)
% (N) / Mean (SD)
% (N) / Mean (SD)
% (N) / Mean (SD)
Demographics
Age, years (range=24-84)
51.6 (13.5)
52.5 (13.7)
50.7 (13.3)
Education, years (4-31)
15.1 (3.4)
15.2 (3.2)
15.0 (3.5)
Gender (% women)
68.9 (91)
65.7 (44)
72.3 (47)
Race (% white)
53.0 (70)
58.2 (39)
47.7 (31)
Psychosocial characteristics
Beck Depression Inventory, score (10-42)
25.3 (8.5)
25.8 (9.2)
24.8 (7.6)
Social support, score (12-31)
22.5 (4.2)
22.2 (4.1)
22.8 (4.2)
Baseline antidepressant treatment (%, n)
34.8 (46)
37.3 (25)
32.3 (21)
Physical health
Physical functioning, score (17-36)
28.9 (5.7)
29.1 (5.6)
28.7 (5.9)
Severity of illness, score (0-22)
6.8 (5.2)
6.5 (4.7)
7.1 (5.7)
Medical diagnoses (%, n)
Peripheral vascular disease
14.4 (19)
14.9 (10)
13.9 ( 9)
Heart/circulatory (other)
13.6 (18)
17.9 (12)
9.2 ( 6)
Gastrointestinal (other)
24.2 (32)
29.9 (20)
9.2 ( 6)
Diabetes (no complications)
15.2 (20)
17.9 (12)
12.3 ( 8)
Musculoskeletal disorders
43.9 (58)
47.8 (32)
40.0 (26)
Urogenital diseases
13.6 (18)
14.9 (10)
12.3 ( 8)
Religious involvement
Affiliation (% Christian)
87.9 (116)
92.5 (62)
83.1 (54)
Importance of religion (% very)
47.0 (62)
44.8 (30)
49.2 (32)
Religious attendance, score (1-6)
3.8 (1.6)
3.9 (1.6)
3.8 (1.6)
Prayer/scripture read, score (1-6)
3.6 (1.7)
3.6 (1.8)
3.6 (1.6)
Intrinsic religiosity, score (12-50)
34.8 (8.4)
34.5 (8.3)
35.2 (8.4)
Daily spiritual experiences, score (19-94)
57.6 (15.9)
57.5 (16.1)
57.7 (15.9)
Religious coping, score (13-42)
29.3 (6.2)
29.1 (5.4)
29.5 (6.9)
Overall religiosity, score (60-190)
129.0 (29.6)
128.6 (29.4)
129.4 (30.1)
Genetic polymorphisms (n=120-128)
5-HTTLPR
L/L (%)
23.2 (29)
15.4 (10)
31.7 (19) *
S/L
57.6 (72)
61.5 (40)
53.3(32)
S/S
21.9 (24)
23.1 (15)
15.0 (9)
5-HTTLPR
LAor LA/LG
79.5 (97)
82.5 (52)
76.3 (45)
(rs25531)
LG
21.3 (25)
17.5 (11)
23.7 (14)
HTR1A
(rs6295)
C/C
21.9 (28)
20.0 (13)
23.8 (15)
G/C
57.0 (73)
60.0 (39)
54.0 (34)
G/G
21.1 (27)
20.0 (13)
22.2 (14)
MAOAuVNTR activity
Low (3/3)
25.6 (32)
26.6 (17)
24.6 (15)
High (any hi risk allele)
74.4 (93)
73.4 (47)
75.4(46)
Any HRG1
Absent
12.5 (15)
12.9 (8)
12.1 (7)
Present
87.5 (105)
87.1 (54)
87.9(51)
1HRG= high risk genotype= S/S, LG, G/G, or high activity MAOA-uVNTR (3.5, 4, or 5)
*p<0.05
Table 1: Characteristics of overall sample and treatment groups.
Religious involvement and genotype
Average levels of religious activity, belief, and experience by genotype are displayed in Table 2. No significant association was found in the overall sample between individual religious measures or overall religiosity and genotype, with two exceptions. There was a tendency for individuals with uMAOA-VNTR high activity alleles to attend religious services less often than those with no high activity allele (3.7 [SD=1.7] vs. 4.3 [SD=1.3], df=123, t=1.76, p=0.08); likewise, those with one or more high risk genotype in the combined analysis attended religious services less frequently than those with no high risk genotype (3.7 [SD=1.7] vs. 4.7 [SD=1.2], df=118, t=2.07, p=0.04).
![]()
Religious Attendance
Prayer/
Scripture
Intrinsic Religiosity
Spiritual Experiences
Religious Coping
Overall Religiosity
Genetic Polymorphism
Mean (SD)
Mean (SD)
Mean (SD)
Mean (SD)
Mean (SD)
Mean (SD)
5-HTTLPR
L/L (n=29)
4.0 (1.5)
3.9 (1.8)
36.2 (8.0)
58.6 (17.3)
29.9 (7.0)
132.7(32.6)
L/S (n=72)
3.7 (1.6)
3.5 (1.7)
35.0 (8.0)
58.6 (15.7)
29.7 (6.1)
130.5(28.7)
S/S (n=24)
3.9 (1.8)
3.5 (1.8)
32.3 (9.9)
53.6 (15.8)
27.3 (5.9)
130.5(28.7)
5-HTTLPR (rs25531)
LA or LA/LG (n=97)
3.9 (1.7)
3.6 (1.7)
34.4 (8.8)
57.0 (16.1)
29.1 (6.3)
127.9(30.5)
LG (n=25)
3.5 (1.3)
3.8 (1.7)
35.4 (6.7)
59.3 (16.1)
29.8 (6.2)
131.8(27.8)
HTR1A (rs6295)
C/C (n=28)
3.9 (1.8)
3.4 (1.9)
36.3 (8.6)
58.6 (15.3)
29.7 (6.6)
132.0(29.3)
C/G (n=73)
3.8 (1.5)
3.6 (1.6)
34.2 (8.2)
56.2 (17.0)
28.8 (6.3)
126.2(30.6)
G/G (n=27)
4.0 (1.7)
4.0 (1.7)
35.3 (8.8)
61.0 (14.0)
30.4 (5.6)
134.7(28.3)
MAOA-uVNTR1
Low (n=32)
4.3 (1.3) #
3.6 (1.8)
34.4 (9.2)
58.3 (18.3)
29.1 (7.2)
128.9(33.8)
High (n=93)
3.7 (1.7)
3.6 (1.7)
35.0 (8.2)
57.2 (15.3)
29.4 (6.0)
128.8(28.7)
Combined2
No HRG3(n=15)
4.7 (1.2) *
4.1 (1.5)
36.6 (8.7)
62.3 (13.2)
31.2 (6.0)
138.8(26.3)
Any HRG (n=105)
3.7 (1.7)
3.6 (1.7)
34.3 (8.4)
56.6 (16.4)
29.0 (6.3)
127.1(30.4)
SD=Standard Deviation
*p<0.05, # 0.05<p<0.10 (analysis of variance or t-test) (n=119-128)
1Low activity=two low activity alleles only (3/3); high activity= any high activity alleles (3.5, 4, or 5)
2Combined= S/S or LG or G/G or any high activity MAOA-uVNTR alleles (3.5, 4, or 5) (high risk genotypes)
3HRG= high risk genotype
Table 2: Average level of religiosity by polymorphism type or combination of polymorphism type (overall sample).
Race
Post-hoc analyses stratified by race focused on Blacks and white Caucasians (analyses not shown). Among Blacks, those with the LG genotypes of the rs25531 (n=9) were less likely to attend religious services than those who were homozygous for the high activity LA alleles (LA/LA, n=12) (3.3 [SD=1.4] vs. 4.8 [SD=1.1], df=19, t=2.72, p=0.01). Daily spiritual experiences were also less frequent among Blacks carrying the LG allele than in those who were homozygous for the LA alleles (54.9 [SD=16.1] vs. 68.3 [SD=13.0], df=19, t=2.12, p<0.05). As a result, overall religiosity tended to be lower among Blacks with the LG genotype than in those who were homozygous for the LA allele (129.3 [SD=30.0] vs. 150.2 [SD=21.4], df=19, t=1.86, p=0.08) (Figure 1).
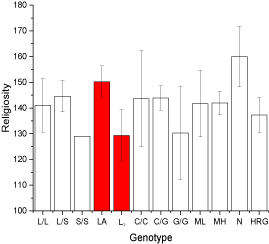
Figure 1: Average overall religiosity (standard error mean) by genotype in
Blacks (n=20-21).
LA=LA/LA, ML=MAOA low activity genotype (3/3), MH=MAOA high activity
genotypes (3.5, 4, or 5); N=no high risk genotypes, HRG=one or more high
risk genotypes (i.e., S/S, LG, G/G, or MH). Sample size ranges from 20-21 due
to incomplete genotype data on one Black participant.
Finally, Blacks with one or more of the four high risk genotypes (i.e., S/S, LG, G/G, or a high activity allele of MAOA-uVNTR [3.5, 4, or 5]) were less likely to attend religious services than those without any high risk genotypes (3.8 [SD=1.4] vs. 5.8 [SD=0.5], df=18, t=2.72, p=0.01), tended to pray, meditate, or read religious scriptures less often (3.8 [SD=1.5] vs. 5.3 [SD=0.50], df=18, t=1.90, p=0.07) and had fewer daily spiritual experiences (59.8 [SD=15.3] vs. 76.0 [SD=12.2], df=18, t=1.97, p=0.07). No other associations in Blacks were significant (or trend), nor were any associations in white Caucasians.
Gender
Post-hoc analyses stratified by gender (analyses not shown) revealed that among men, those with any high risk genotypes in the combined analysis were less likely to report daily spiritual experiences (51.3 [SD=17.6] vs. 65.3 [SD=13.4], df=33, t=2.06, p=0.047) and scored lower on overall religiosity (117.5 [SD=30.8] vs. 144.5 [SD=28.3], df=33, t=2.21, p=0.03) compared to those with no high risk genotypes. No other associations in men were statistically significant, nor were any associations in women.
Effect of genotype on treatment response
Trajectories of response to RCBT (1) vs. CCBT (0) in each genotype were examined using mixed effect regression models (Table 3). Each model included the fixed effects of treatment group, time, time squared, and the interaction of treatment group with time (as described in the primary report from the trial [51]). In the overall sample, there were no significant differences in treatment response to RCBT vs. CCBT, except in those with the C/C genotype of the HTR1A (rs6295), where RCBT was particularly effective in reducing depressive symptoms compared to CCBT (group x time interaction B=3.33, SE=1.17, df=62, t=2.86, p=0.0058, n=28, Cohen’s d=0.73) (Figure 2). When analyses were stratified by overall religiosity, effects were particularly strong in those with low religiosity (group x time interaction B=4.32, SE=1.56, p=0.0097). Those with no high risk genotype were also more likely to respond to RCBT (group x time interaction B=3.40, SE=1.60, df=32, t=2.12, p=0.042, n=15, d=0.75).
![]()
Response to RCBT vs. CCBT
Genetic Polymorphism
Overall
Low Religiosity
High Religiosity
5-HTTLPR
Estimate (SE) 1
Estimate (SE)
Estimate (SE)
L/L (n=29)
-1.97 (3.51)
-5.20 (5.12)
-0.56 (4.71)
S/L (n=72)
1.49 (2.58)
3.78 (3.60)
-1.13 (3.76)
S/S (n=24)
-1.11 (4.36)
1.03 (5.16)
-7.33 (7.33)
5-HTTLPR (rs25531)
LA or LA/LG (n=97)
0.11 (2.05)
1.08 (2.92)
-1.05 (2.82)
LG (n=25)
1.75 (4.80)
4.33 (6.48)
-1.69 (7.24)
HTR1A (rs6295)
C/C (n=28)
-5.83 (3.66)
-6.67 (5.02)
-6.44 (5.54)
C/G (n=73)
3.43 (2.43)
6.99 (3.24)*
-0.40 (3.63)
G/G (n=27)
-1.31 (4.15)
-3.80 (7.08)
0.93 (5.18)
MAOA-uVNTR2
Low (n=32)
-4.29 (3.55)
1.36 (4.48)
-9.42 (5.94)
High (n=93)
2.16 (2.16)
3.37 (3.11)
1.07 (2.95)
Combined3
No HRG (n=15)
-5.28 (5.24)
7.53 (9.26)
-4.61 (7.66)
Any HRG4 (n=105)
1.38 (2.05)
2.96 (2.76)
-0.46 (3.04)
*p<0.05
RCBT = religious-integrated cognitive behavioral therapy; CCBT = conventional CBT
1Unstandardized beta (B or estimate) and standard error (SE) from mixed model (intent-to-treat analysis at 12 weeks), where RCBT=1 and CCBT=0
2Low activity= two low activity alleles only (3/3); high activity= any high activity alleles (3.5, 4, or 5)
3Combined=S/S or LG or G/G or any high activity MAOA-uVNTR alleles (3.5, 4, or 5) (high risk genotypes)
4HRG= high risk genotype
Table 3: Treatment response to RCBT vs. CCBT by polymorphism type or combination of polymorphism type for overall sample and for sample divided by level of religiosity.
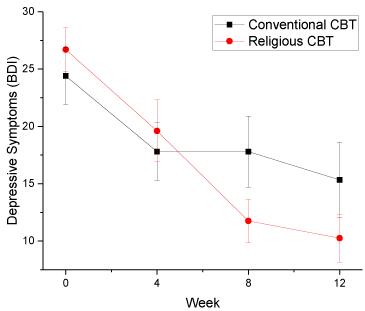
Figure 2: Changes in Beck Depression Inventory (BDI) in response to
religious vs. conventional cognitive behavioral therapy in those with HTR1A
genotype C/C (standard error bars) (n=28).
Discussion
This is one of the first studies to examine the relationship between religious involvement and candidate polymorphisms at the serotonin transporter gene, serotonin 1A receptor gene, and MAOA gene. To our knowledge, this is the first study to examine those relationships in persons with major depressive disorder in the setting of chronic medical illness (a common stressor for many). This is also the first study to examine the effects of genotype on trajectories of response to religious vs. conventional CBT. Although the results presented here are highly exploratory in nature, the analyses were hypothesis driven and plausible given the existing research. However, none of our three hypotheses was confirmed, namely religiosity was not associated with most genotypes conveying high risk for depression; the effectiveness of the two types of therapy did not vary based on the patient’s genotype, except in those with the C/C genotype of the HTR1A gene and those with no high risk genotype; and RCBT was not more effective in participants with high risk genotypes who were more religious than other participants.
Few relationships were found between religious activities, attitudes, or experiences and genotypes examined in this study. The only significant finding in the overall sample was that religious attendance was less common among those with at least one high risk genotype, and tended to be less common in those with high activity alleles of the MAOA-uVNTR gene. In post-hoc analyses that involved stratification by race and gender, several additional significant relationships were found, again in the opposite direction expected. Among Blacks, the presence of one or more high risk genotype, especially those containing the low functioning LG genotype of the rs25531 polymorphism (SLC6A4 gene), was associated with less frequent attendance at religious services and fewer daily spiritual experiences. A similar relationship was found in men, where the presence of one or more high risk genotype was associated with fewer daily spiritual experiences and lower overall religiosity.
Thus, while there were few findings, they were at least consistent. If replicated in future studies, this could help to explain why greater religious involvement is associated with lower rates of depression and perhaps faster recovery from depression in those with chronic medical illness, at least among Blacks and men. Those who develop depression may be at greater risk of having certain genotypes that both increase their risk of depression and lower the likelihood of religious activity, thus resulting in an inverse relationship between religiosity and depression in this setting. Admittedly, this explanation is highly speculative, based on the results of only a single study -- a study in which neither religiosity nor genotype were related to depressive symptoms. However, it is a plausible explanation that could help to account for relationships previously reported between religiosity and depression. The findings reported here are also consistent with earlier research in 2,537 healthy community-dwelling adolescents and young adults (one of the few studies, if not only study reporting associations between religious involvement and genotype), where presence of the high risk S allele of 5-HTTLPR was associated with lower religious involvement, a finding that was particularly strong in Blacks [65].
Although we hypothesized that religious CBT would be more effective than conventional CBT in those with high risk genotypes, we actually found the opposite with regard to the HTR1A polymorphism. RCBT was more effective than CCBT in those with the low risk C/C genotype, especially in those with low religiosity. Furthermore, RCBT was also more effective in those without any high risk genotype. Neither of these findings are consistent with our hypothesis. They are consistent, however, with the finding of higher religiosity in those with low risk genotypes. Thus, religiosity is both higher in those with low risk genotypes (at least in certain subgroups) and religious CBT is also more effective than conventional CBT in depressed persons with the low risk C/C genotype and in those without any high risk genotype.
Limitations
Several aspects of the sample, methodology, and analysis limit the generalizability of these results and the conclusions that can be drawn from them. First, this was a population recruited for a treatment trial of depression (rather than an epidemiological sample with a defined sampling frame) and religion/spirituality was at least somewhat important for all participants and all had major depressive disorder and chronic medical illness. The prevalence of the S allele among white Caucasians and Blacks in our sample (79% and 45%, respectively) was considerably higher than others have reported in non-depressed healthy community samples (50% and 30%) [20], and although the prevalence of the LG allele in white Caucasians here (13%) was similar to that reported by others (8%), it was much higher in Blacks in our sample (43%) compared to that reported by others (21%) [21]. Second, our relatively small sample size limited the power to identify significant associations, especially in subgroup analyses stratified by race and gender. Studies examining psychosocial correlates of genotype typically require thousands of subjects. Third, there is controversy in the literature over which genotypes if any are considered high risk for the development of depression. Nevertheless, there is considerable literature to support the particular high risk genotypes chosen here, and the proteins they code for are known to affect levels of neurotransmitters in the brain that influence mood states. Finally, the multiple statistical tests performed in this report make it likely that several of the significant findings reported were due to chance alone. Using the conservative Bonferroni method to correct for multiple comparisons would have reduced the level of statistical significance to p<0.0005. However, given the exploratory nature of these analyses we felt that using the traditional definition of statistical significance (p<0.05) for reporting these initial findings was justified.
Conclusion
We found minimal evidence that religious involvement is associated with a genotype associated with increased risk of depression, although that evidence was consistent. In the overall sample, participants with one or more high risk genotypes were less likely to attend religious services. In subgroup analyses, Blacks with high risk genotypes were less likely to engage in religious activities or have daily spiritual experiences, and men with such genotypes were less likely to have spiritual experiences and were less religious overall. Finally, individuals with the HTR1A genotype C/C (especially those who were low on religiosity) had a more vigorous response to religious CBT than to conventional CBT, as did participants with no high risk genotype, which could have treatment implications. Whether these findings reflect true associations and effects or are the result of multiple statistical comparisons or the unique nature of this sample is unknown. However, they suggest worthy areas to research in different locations and larger samples.
References
- Silveira LA. Experimenting with spirituality: analyzing The God Gene in a nonmajors laboratory course. CBE Life Sci Educ. 2008; 7: 132-145.
- Rowthorn R. Religion, fertility and genes: a dual inheritance model. Proc Biol Sci. 2011; 278: 2519-2527.
- Koenig JB, Bouchard TJ. Genetic and environmental influences on the traditional moral values triad -- authoritarianism, conservatism and religiousness. McNamara P, editor. In: Where God and Science Meet, volume 1. Westport, CT: Praeger, 2006; 31-60.
- Sasaki JY, Kim HS, Xu J. Religion and well-being: The moderating role of culture and oxytocin receptor (OXTR) gene. Journal of Cross-Cultural Psychology. 2011; 42: 1394-1405.
- Rodrigues SM, Saslow LR, Garcia N, John OP, Keltner D. Oxytocin receptor genetic variation relates to empathy and stress reactivity in humans. Proc Natl Acad Sci USA. 2009; 106: 21437-21441.
- Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S, Benjamin J. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science. 1996; 274: 1527-1531.
- Hu XZ, Lipsky RH, Zhu G, Akhtar LA, Taubman J, Greenberg BD, et al. Serotonin transporter promoter gain-of-function genotypes are linked to obsessive-compulsive disorder. American Journal of Human Genetics. 2006; 78: 815-826.
- Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H, et al. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science. 2003; 301: 386-389.
- Risch N, Herrell R, Lehner T, Liang KY, Eaves L, Hoh J, et al. Interaction between the serotonin transporter gene (5-HTTLPR), stressful life events, and risk of depression: a meta-analysis. JAMA. 2009; 301: 2462-2471.
- Rutter M, Thapar A, Pickles A. Gene-environment interactions: biologically valid pathway or artifact? Arch Gen Psychiatry. 2009; 66: 1287-1289.
- Culverhouse RC, Bowes L, Breslau N, Nurnberger JI, Burmeister M, Fergusson DM, et al. Protocol for a collaborative meta-analysis of 5-HTTLPR, stress, and depression. BMC Psychiatry. 2013; 13: 304.
- Cervilla JA, Rivera M, Molina E, Torres-González F, Bellón JA, Moreno-Küstner B, et al. The 5-HTTLPR s/s polymorphism at the serotonin transporter gene (SLC6A4) increases the risk for depression in a large cohort of primary care attendees: The PREDICT-Gene Study. American Journal of Medical Genetics Part B Neuropsychiatric Genetics. 2006; 141B: 912–917.
- Cervilla JA, Molina E, Rivera M, Torres-Gonzalez F, Bellon JA, Moreno B, et al. The risk for depression conferred by stressful life events is modified by variation at the serotonin transporter 5HTTPLPR genotype – evidence for the Spanish PREDICT-gene cohort. Molecular Psychiatry. 2007; 12: 748-755.
- Nakatani D, Sato H, Sakata Y, Shiotani I, Kinjo K, Mizuno H, et al. Influence of serotonin transporter gene polymorphism on depressive symptoms and new cardiac events after acute myocardial infarction. American Heart Journal. 2005; 150: 652-658.
- Otte C, McCaffery J, Ali S, Whooley MA. Association of a serotonin transporter polymorphism (5-HTTLPR) with depression, perceived stress, and norepinephrine in patients with coronary disease: the Heart and Soul Study. Am J Psychiatry. 2007; 164: 1379-1384.
- Zalsman G, Huang YY, Oquendo MA, Burke AK, Hu XZ, Brent DA, et al. Association of a triallelic serotonin transporter gene promoter region (5- HTTLPR) polymorphism with stressful life events and severity of depression. American Journal of Psychiatry. 2006; 163: 1588–1593.
- Wang Z, Baker DG, Harrer J, Hammer M, Price M, Amstadter A. Relationship between combat-related posttraumatic stress disorder and the 5-HTTLPR/rs25531 polymorphism. Depression & Anxiety. 2011; 28: 1067-1073.
- Lotrich FE, Ferrell RE, Rabinovitz M, Pollock BG. Risk for depression during interferon-alpha treatment is affected by the serotonin transporter polymorphism. Biological Psychiatry. 2009; 65: 344–348.
- Mak KK, Kong WY, Mak A, Sharma VK, Ho RC. Polymorphisms of the serotonin transporter gene and post-stroke depression: a meta-analysis. J Neurol Neurosurg Psychiatry. 2013; 84: 322-328.
- Gelemter. American Journal of Genetics. Neuropsychiatr Genet. 2009; 88: 61-66.
- Odgerel Z, Talati A, Hamilton SP, Levinson DF, Weissman MM. Genotyping serotonin transporter polymorphisms 5-HTTLPR and rs25531 in European- and African-American subjects from the National Institute of Mental Health's Collaborative Center for Genomic Studies. Transl Psychiatry. 2013; 3: e307.
- Williams RB, Marchuk DA, Gadde KM, Barefoot JC, Grichnik K, Helms MJ, et al. Serotoni-related gene polymorphisms and central nervous system serotonin function. Neuropsychopharmacology. 2003; 28: 533-541.
- Brummett BH, Boyle SH, Siegler IC, Kuh CM, ashley-Koch a, Jonassaint CR, et al. Effects of environmental stress and gender on associations among symptoms of depression and the serotonin transporter gene linked polymorphic region (5-HTTLPR). Behavior Genetics.2008; 38: 34-43.
- Rivera M, Molina E, Torres-González F, Bellón J, Moreno-Küstner B, King M, et al. High-activity variants of the uMAOA polymorphism increase the risk for depression in a large primary care sample. American Journal of Medical Genetics. Part B: Neuropsychiatric Genetics. 2009; 5: 395-402.
- Deckert J, Catalano M, Syagailo YV, Bosi M, Okladnova O, Di Bella D, et al. Excess of high activity monoamine oxidase A gene promoter alleles in female patients with panic disorder. Hum Mol Genet. 1999; 8: 621–624.
- Denney RM, Koch H, Craig IW. Association between monoamine oxidase A activity in human male skin fibroblasts and genotype of the MAOA promoter-associated variable number tandem repeat. Hum Genet. 1999; 105: 542-551.
- Schulze TG, Müller DJ, Krauss H, Scherk H, Ohlraun S, Syagailo YV, et al. Association between a functional polymorphism in the monoamine oxidase A gene promoter and major depressive disorder. Am J Med Genet. 2000; 96: 801-803.
- Yu YW, Tsai SJ, Hong CJ, Chen TJ, Chen MC, Yang CW. Association study of a monoamine oxidase a gene promoter polymorphism with major depressive disorder and antidepressant response. Neuropsychopharmacology. 2005; 30: 1719-1723.
- Eley TC, Tahir E, Angleitner A, Harriss K, McClay J, Plomin R, et al. Association analysis of MAOA and COMT with neuroticism assessed by peers. Am J Med Genet B Neuropsychiatr Genet. 2003; 120B: 90-96.
- Parsey RV, Oquendo MA, Ogden RT, Olvet DM, Simpson N, Huang YY, et al. Altered serotonin 1A binding in major depression: a [carbonyl-C-11]WAY100635 positron emission tomography study. Biological Psychiatry. 2006; 59: 106–113.
- Lemonde S, Turecki G, Bakish D, Du L, Hrdina PD, Bown CD, et al. Impaired repression at a 5-hydroxytryptamine 1A receptor gene polymorphism associated with major depression and suicide. J Neurosci. 2003; 23: 8788-8799.
- Koenig HG, George LK, Peterson BL. Religiosity and remission of depression in medically ill older patients. Am J Psychiatry. 1998; 155: 536-542.
- Koenig HG. Religion and remission of depression in medical inpatients with heart failure/pulmonary disease. J Nerv Ment Dis. 2007; 195: 389-395.
- Bellamy CD, Jarrett NC, Mowbray O, MacFarlane P, Mowbray CT, Holter MC. Relevance of spirituality for people with mental illness attending consumer-centered services. Psychiatr Rehabil J. 2007; 30: 287-294.
- Vaillant G, Templeton J, Ardelt M, Meyer SE. The natural history of male mental health: health and religious involvement. Soc Sci Med. 2008; 66: 221-231.
- Freud, S. Future of an Illusion. Strachey J editor and translator. In: Standard Edition of the Complete Psychological Works of Sigmund Freud. London: Hogarth Press. 1962; 43.
- Andrews PW, Thomson JA. Depression’s evolutionary roots. Scientific American. 2009.
- De Catanzaro D. Suicidal ideation and the residual capacity to promote inclusive fitness: a survey. Suicide Life Threat Behav. 1984; 14: 75-87.
- Koenig HG, King DE, Carson VB. Handbook of Religion and Health. NY, NY: Oxford University Press. 2012; 145-173.
- Bachner-Melman R, Dina C, Zohar AH, Constantini N, Lerer E, Hoch S, et al. AVPR1a and SLC6A4 gene polymorphisms are associated with creative dance performance. PLoS Genet. 2005; 1: e42.
- Levin JS, Wickramasekera IE, Hirshberg C. Is religiousness a correlate of absorption? Implications for psychophysiology, coping, and morbidity. Altern Ther Health Med. 1998; 4: 72-76.
- Kim SJ, Lee HS, Kim CH. Obsessive-compulsive disorder, factor-analyzed symptom dimensions and serotonin transporter polymorphism. Neuropsychobiology. 2005; 52: 176-182.
- Bachner-Melman R, Dina C, Zohar AH, Constantini N, Lerer E, Hoch S, et al. AVPR1a and SLC6A4 gene polymorphisms are associated with creative dance performance. PLoS Genet. 2005; 1: e42.
- Lorenzi C, Serretti A, Mandelli L, Tubazio V, Ploia C, Smeraldi E. 5-HT 1A polymorphism and self-transcendence in mood disorders. Am J Med Genet B Neuropsychiatr Genet. 2005; 137B: 33-35.
- Borg J, Andrée B, Soderstrom H, Farde L. The serotonin system and spiritual experiences. Am J Psychiatry. 2003; 160: 1965-1969.
- Steiger H, Joober R, Gauvin L, Bruce KR, Richardson J, Israel M, et al. Serotonin-system polymorphisms (5-HTTLPR and 1438G/A) and responses of patients with bulimic syndromes to multimodal treatments. Journal of Clinical Psychiatry. 2008; 69: 1565-1571.
- Schosser A, Kasper S. The role of pharmacogenetics in the treatment of depression and anxiety disorders. Int Clin Psychopharmacol. 2009; 24: 277-288.
- Dreimuller N, Tadic A, Dragicevic A, Boland K, Bondy B, Lieb K, et al. The serotonin transporter promoter polymorphism (5-HTTLPR) affects the relation between antidepressant serum concentrations and effectiveness in major depression. Pharmacopsychiatry. 2012; 45: 108-113.
- Hayden EP, Dougherty LR, Maloney B, Olino TM, Sheikh H, Durbin CE, et al. Early-emerging cognitive vulnerability to depression and the serotonin transporter promoter region polymorphism. Journal of Affective Disorders. 2008; 107: 227-230.
- Pearce MJ, Koenig HG, Robins CJ, Nelson B, Shaw SF, Cohen HJ, et al. Religiously-integrated cognitive behavioral therapy: A new method of treatment for major depression in patients with chronic medical illness. Psychotherapy. 2015; 52: 56-66.
- Koenig HG, Pearce MJ, Nelson B, Shaw SF, Robins CJ, Daher N, et al. Religious vs. conventional cognitive-behavioral therapy for major depression in persons with chronic medical illness. Journal of Nervous and Mental Disease. 2015; 203: 243-251.
- Koenig HG. An abbreviated Mini-Mental State Exam for medically ill older adults. J Am Geriatr Soc. 1996; 44: 215-216.
- Sheehan BV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, et al. The Mini International Neuropsychiatric Interview (MINI): The development and validation of structured diagnostic psychiatric interview for DSM-IV and ICD-10. Journal of Clinical Psychiatry. 1998; 59: 22–33.
- Beck AT, Ward CH., Mendelson M, Mock J, Erbaugh J. An inventory for measuring depression. Archives of General Psychiatry. 1961; 4: 561-571.
- Hlatky MA, Boineau RE, Higginbotham MB, Lee KL, Mark DB, Califf RM, et al. A brief self-administered questionnaire to determine functional capacity (the Duke Activity Status Index). Am J Cardiol. 1989; 64: 651-654.
- Linn BS, Linn MW, Gurel L. Cumulative illness rating scale. J Am Geriatr Soc. 1968; 16: 622-626.
- Landerman R, George LK, Campbell RT, Blazer DG. Alternative models of the stress buffering hypothesis. Am J Community Psychol. 1989; 17: 625-642.
- Hoge DR. A validated intrinsic religious motivation scale. Journal for the Scientific Study of Religion. 1972; 11: 369-376.
- Underwood LG, Teresi JA. The daily spiritual experiences scale: development, theoretical description, reliability, exploratory factor analysis, and preliminary construct validity using health-related data. Annals of Behavioral Medicine. 2002; 24: 22-33.
- Pargament KI, Smith BW, Koenig HG, Perez L. Patterns of positive and negative religious coping with major life stressors. Journal for the Scientific Study of Religion. 1998; 37: 710-724.
- Koenig HG. Religious vs. conventional psychotherapy for major depression in patients with chronic medical illness: Rationale, methods, and preliminary results. Depression Research & Treatment Article. 2012.
- Beck AT, Rush J, Shaw BF, Emery G. Cognitive Therapy of Depression. NY, NY: Guilford Press. 1979.
- Beck JS. Cognitive Therapy for Challenging Problems. NY, NY: Guilford Press. 2005.
- Hu X, Oroszi G, Chun J, Smith TL, Goldman D, Schuckit MA. An expanded evaluation of the relationship of four alleles to the level of response to alcohol and the alcoholism risk. Alcohol Clin Exp Res. 2005; 29: 8-16.
- Dew RE, Koenig HG. Religious involvement, the serotonin transporter promoter polymorphism, and drug use in young adults. International Journal of Soccial Science Studies. 2014; 2: 98-104.